Anatomie des insectes

Anatomie d'une mouche tsé-tsé : en violet, la tête ; en bleu, le thorax ; en vert, l'abdomen
L’anatomie des insectes qui décrit la morphologie de leurs structures externes et internes, et les principales propriétés de ces structures, présente de nombreux points communs avec celle des Arthropodes du sous-embranchement des Hexapodes : le corps est segmenté (d'où le nom d'insecte qui est un emprunt savant — daté de 1553 — au latin insectus, pluriel neutre substantivé de insectus, participe passé de insecare, « couper »)[1], les biologistes appelant ces segments des métamères ; l'altération fonctionnelle de la métamérie, appelée tagmatisation, en lien avec l'adaptation aux milieux de vie très diversifiés (l’adaptation des différents membres d'une guilde écologique à des espaces écologiques, des modes de vie, implique les tagmes qui acquièrent une spécialisation morphologique et fonctionnelle plus ou moins poussée), se traduit par un corps divisé en trois tagmes céphalique, thoracique et abdominal. La tête est spécialisée dans la fonction sensorielle et trophique avec notamment une paire d'antennes, de yeux composés et des pièces buccales, un thorax à fonction essentiellement locomotrice grâce à trois paires de pattes articulées, et un abdomen, dépourvu d'appendices autres que génitaux, qui a une fonction de digestion, d'excrétion et de reproduction. Les pièces buccales sont visibles et externes (avec notamment les mandibules, mâchoires capables de mordre),, d'où l'appellation ancienne d'Ectognathes ou insectes vrais, ce qui les distinguent des Entognathes, petite classe d'hexapodes (Collemboles, Diploures, Protoures), vraisemblablement paraphylétique, aux pièces buccales internes non apparentes.
Le corps des insectes, comme celui des autres arthropodes, est recouvert d'un exosquelette rigide composé d'une molécule caractéristique, la chitine, et constitué de trois couches de cuticule.
Sommaire
1 Anatomie externe
1.1 Tête
1.1.1 Ocelles
1.1.2 Yeux composés
1.1.3 Antennes
1.1.3.1 Types communs d'antennes d'insectes
1.1.4 Pièces buccales
1.1.4.1 Suceur simple
1.1.4.2 Suceur-piqueur
1.1.4.3 Suceur-spongieur
1.1.4.4 Suceur-lécheur
1.1.4.5 Galerie pièces buccales
1.2 Thorax
1.2.1 Pattes
1.2.2 Ailes
1.3 Abdomen
1.4 Exosquelette
2 Anatomie interne
2.1 Système respiratoire
2.2 Système circulatoire
2.3 Système digestif
2.4 Système nerveux central
2.5 Système reproducteur
2.5.1 Femelle
2.5.2 Mâle
3 Schéma anatomie interne
4 Références
5 Voir aussi
Anatomie externe |
Comme tous les arthropodes, les insectes ont un corps segmenté soutenu par un exosquelette qui est composé d'une cuticule chitineuse. Les segments du corps sont organisés en trois parties principales qui sont la tête, le thorax et l'abdomen. La tête possède une paire d'antennes, une paire d'yeux composés, des ocelles et trois ensembles d'appendices modifiés qui forment les pièces buccales.

Tête chez l'insecte
Le thorax est composé de trois segments (prothorax, mésothorax et le métathorax) et porte généralement tous les organes locomoteurs (ailes ou pattes). L'abdomen est composé la plupart du temps de onze segments qui peuvent parfois porter des appendices tels des cerques par exemple. À l'intérieur, il contient une partie des organes importants comme l'appareil digestif, le système respiratoire, le système excréteur et les organes reproducteurs[2]. On retrouve une grande variabilité et de nombreuses adaptations dans la physiologie des parties du corps de l'insecte, en particulier les ailes, les pattes, les antennes et les pièces buccales.
Tête |
Chez la plupart des insectes au stade adulte ou larvaire, la tête est entourée d'un disque fortement sclérifié que l'on nomme capsule céphalique ou épicrâne. Cette formation n'est cependant pas présente chez toutes les larves d'insectes (ex: larves de mouche Cyclorrhapha). La capsule céphalique est la partie principale de la tête et elle porte les yeux composés, les ocelles, une paire d'antennes et les pièces buccales. Cette capsule est l'assemblage de 6 segments les plus antérieurs du corps[3].
L'orientation de l'espace préoral (délimité par les pièces buccales) permet de distinguer la tête orthognathe (l'ouverture de l'espace préoral est perpendiculaire à l'axe du corps), la tête prognathe (l'ouverture s'est déplacée vers l'avant), et la tête opisthognathe (l'ouverture s'est déplacée vers l'arrière)[4].
Le frons est la partie de la capsule céphalique qui se retrouve à l'avant, juste en dessous des antennes. Il varie en forme et en taille. Chez certaines espèces, sa délimitation est relativement arbitraire. Chez d'autres, il est délimité par des sillons fronto-clypéaux ou épistomiaux. Dans le cas des insectes à trois ocelles, le médian est habituellement situé sur le frons. Sur le sommet de la tête (vertex), on retrouve des sutures frontales qui peuvent être de différentes formes (Y, U ou V). Lors de la mue de certains insectes, ces sutures s'ouvrent et permettent à l'animal de s'extirper de son ancienne enveloppe. Les sutures frontales ne sont pas présentes chez tous les insectes.
Les quatre segments les plus apparents sont ceux qui ont conservé des appendices pairs articulés : antennes, mandibules, maxilles et labium.
Ocelles |
Les ocelles sont des organes photorécepteurs simples qui se retrouvent sur le dessus de la tête des insectes. Ces organes sont extrêmement sensibles à la lumière mais leur fonction n'est pas visuelle. Ils sont généralement au nombre de trois, un central et deux latéraux. Certains insectes terrestres, comme certaines fourmis et cafards, n'ont que deux ocelles. Les ocelles détectent la luminosité ainsi que les petites variations dans celle-ci.
Les ocelles pourraient aider l'insecte à maintenir la stabilité pendant le vol grâce à cette capacité à détecter les changements dans la luminosité[5],[6] . Ils joueraient également un rôle dans la visualisation de la polarité ou encore comme détecteur du cycle circadien.
Yeux composés |

Yeux composés (photo prise au microscope)
Les yeux composés sont formés par la juxtaposition de plusieurs centaines d'ommatidies (jusqu'à 30 000 dans un œil composé unique), une unité visuelle simple. L'image perçue est donc une combinaison des contributions de ses nombreuses ommatidies. Comparativement aux yeux simples, ils ont moins de résolution mais ils possèdent un plus grand angle de vue et sont capables de détecter les mouvements rapides. Dans certains cas, ils ont la possibilité de détecter la polarité[7].

Pseudopupille observable chez la mante.
Chez les insectes volants ou prédateurs, les ommatidies sont organisées de manière à donner une vision très spécialisée. Les unités visuelles sont plus aplaties et leurs facettes sont beaucoup plus grandes. L'aplatissement permet aux ommatidies de recevoir une plus grande quantité de lumière. La résolution des images se retrouve donc plus élevée.
Chez ces insectes, on retrouve la pseudopupille (en), une petite tache noire qui semble toujours regarder en direction de l'observateur. Cette tache résulte de l'absorption de la lumière par le rhabdome des ommatidies dont l'axe coïncide avec l'axe de l'observateur, tandis que les ommatidies de côté la reflètent[8].
Antennes |
Les antennes sont les principaux organes de l'odorat. Elles peuvent détecter les mouvements et leur orientation, les odeurs, les sons, l'humidité et une variété de signaux chimiques (phéromones). Elles peuvent également servir d'organes tactiles pour analyser l'environnement ; de moyen de communication pour l'accouplement ou encore de moyen de défense. L'antenne est composée de trois sections principales : le scape, le pédicelle et le flagelle.
Scape : le premier article (le plus basal) de l'antenne.
Pédicelle : le deuxième article de l'antenne.
Flagelle : l'ensemble des articles de l'antenne à partir du troisième article.
Flagomères : les articles qui composent la flagelle.
Les antennes sont très variables chez les insectes[9]. De plus, au sein de la même espèce, on retrouve également de légères différences entre les sexes.
Types communs d'antennes d'insectes |

Différents types d'antennes d'insectes
Aristée : forme élargie avec un poil latéral (ex : Diptera).
Filiforme : forme simple, allongée et droite.
Sétacée : l'antenne se rétrécit progressivement de la base à la pointe (ex : Thysanoptera, Blattaria, Ephemeroptera, Plecoptera et Trichoptera).
Moniliforme : segments antennaires en forme ronde qui donnent une apparence de collier de perles (ex : Coleoptera).
Serriforme : antenne qui est inclinée d'un côté donnant l'apparence d'un bord de scie (ex : Coleoptera).
Pectiniforme : antenne ayant l'apparence d'un peigne, les segments sont plus longs d'un côté (ex : Coleoptera, Hymenoptera - Symphyta).
Claviforme : segments antennaires qui s'élargissent à la pointe de l'antenne. Cela peut être progressif et présent sur toute la longueur ou une augmentation soudaine dans les derniers segments (ex : Coleoptera et Lepidoptera).
Lamelliforme : segments antennaires aplatis formant des lamelles (ex : Coleoptera - Scarabeidae).
Coudée ou géniculée : présence d'un coude dans l'antenne (Hymenoptera - Formicidae et Coleoptera).
Plumeuse : Segments antennaires avec un certain nombre de branches fines, semblable à une plume (ex : Diptera et Lepidoptera - Saturniidae)
Pièces buccales |

Pièces buccales : .mw-parser-output .sep-liste{font-weight:bold}
A. broyeur (ex: criquet), B. suceur-lécheur (ex: abeille), C. suceur (ex: papillon), D. suceur-piqueur (ex: moustique), a. antenne, c. yeux composés, lb. labium, lr. labre, md. mandibules, mx. maxilles, hp. hypopharynx
Les invertébrés ancestraux avaient des pièces buccales du type broyeur. Au cours de l'évolution, une spécialisation s'est effectuée, si bien que maintenant on en retrouve plusieurs types (broyeur, suceur, suceur-piqueur, suceur-spongieur et suceur-lécheur)[10].
Chez les broyeurs, on retrouve quatre pièces d'importance : labre, mandibules, maxilles et le labium. Les trois dernières sont des appendices locomoteurs ancestraux qui se sont modifiés et spécialisés à des fonctions reliées à l'alimentation.
Labre : C'est une pièce unique de la bouche formant la lèvre supérieure. Il couvre la base des mandibules et forme la voûte de la cavité buccale des insectes.
Mandibules : Elles servent à saisir, couper ou encore broyer la nourriture. Elles sont les principaux organes d'ingestion et les premiers appendices buccaux.
Maxilles : Elles sont accessoires aux mandibules et elles peuvent avoir plusieurs fonctions (masticatrices ou sensorielles).
Labium : Il est la fusion de la deuxième paire de maxilles. Il joue un rôle de lèvre inférieure et peut être utilisé comme organe sensoriel.
Les pièces buccales du type broyeur se rencontre chez plusieurs ordres d'insectes (coléoptères, orthoptères, odonates, plécoptères, neuroptères, certains hyménoptères, larves de lépidoptères etc.).
Suceur simple |
Ce type de pièces buccales se rencontre chez les papillons (lépidoptères). Les maxilles ont évolué en une trompe multi segmentée qu'on appelle le proboscis. Au repos, ce tube est enroulé sous la tête. Les papillons se nourrissent du nectar des fleurs, des sels minéraux et des nutriments contenus dans d'autres liquides. On retrouve également deux longs palpes incurvés vers l'avant qui sont dérivés du labium.
Suceur-piqueur |
Plusieurs ordres d'insectes ont des pièces buccales qui percent pour ensuite s'alimenter par succion des fluides internes des plantes ou des organismes vivants. Certains sont phytophages, comme les pucerons et les cicadelles, tandis que d'autres sont insectivores, prédateurs ou hématophages, comme les moustiques ou certaines punaises.
Chez ces insectes, le labium forme un proboscis de quatre sections articulées entre elles. À l'intérieur, on retrouve quatre stylets dont le plus externe est la dérivation des mandibules et le plus interne les maxilles. À l'intérieur, on retrouve un canal d'ingestion et un canal salivaire.
Chez les moustiques femelles, le labium forme une gaine et renferme toutes les autres pièces buccales. Le labre forme un tube qui sert à aspirer le sang. Les mandibules et les maxilles ont évolué en quatre stylets qui servent à percer la peau de la victime.
Suceur-spongieur |
Certaines mouches (ex: mouche domestique) ont des pièces buccales du type suceur-spongieur. Le labium est modifié en forme de trompe et il sert à aspirer les aliments liquides jusqu'à l'œsophage. Au bout de celui-ci, on retrouve des lamelles qui forment un appareil spongieux. Cet organe fonctionne comme une éponge et absorbe les aliments sous forme liquide. Pour manger de la nourriture solide, la mouche sécrète de la salive et la dépose sur les aliments. Sa salive est remplie d'enzymes digestives et dissout la nourriture. Le résultat est ensuite aspiré dans le labium par capillarité.
Suceur-lécheur |
Certains hyménoptères (ex: abeille domestique et bourdons) ont des pièces buccales du type suceur-lécheur. Chez ces insectes, le labium est allongé pour former une sorte de langue ou de glosse. Cette langue imbibe le nectar des fleurs ou encore d'autres liquides et le transfère à l'œsophage. Ils sont également munis de mandibules, qu'ils utilisent pour saisir, couper ou broyer[11].
Galerie pièces buccales |

Broyeur

Suceur simple

Suceur-spongieur

Suceur-lécheur

Suceur-piqueur





Thorax |

Thorax chez l'insecte :
1. Prescutum, 2. Scutum, 3. Scutellum, 4. Propleuron, 5. Mesopleuron, 6. Metapleuron, 7. Prosternum, 8. Mesosternum, 9. Metasternum
Le thorax est situé entre la tête et l'abdomen. Chez la majorité des insectes, il est formé de trois parties distinctes : prothorax (T1), mésothorax (T2) et le métathorax (T3). Ces trois segments sont entourés de sclérites formant l'exosquelette de chaque segment : notum (ou scutum), sur la face dorsale, pleuron, sur les côtés, sternum, sur la face ventrale. Chaque segment décline ces noms : ainsi les sclérites du prothorax sont les pronotum (ou prescutum), propleuron, et prosternum, ceux du mésothorax les mésonotum (ou scutum), mésopleuron, et mésosternum, ceux du métathorax les métanotum (ou scutellu), métapleuron, et métasternum.
C'est la partie du corps à laquelle les trois paires de pattes et les ailes sont attachées. Le thorax est donc parfaitement adapté à la locomotion (marche, nage et vol). Chez les insectes ptérygotes adultes, les segments T2 et T3 portent les ailes.
Pattes |
 Lire le média
Lire le médiaDémarche tripode d'une fourmi (insecte).
Les pattes sont composées de plusieurs segments : coxa, trochanter, fémur, tibia et tarse. Ce dernier est segmenté en plusieurs parties appelées tarsomères. À l'extrémité du tarse on trouve une ou deux griffes. Le fémur et le tibia sont généralement les sections les plus longues.
Les pattes servent principalement à la locomotion. L'évolution des membres locomoteurs des arthropodes est en lien avec leur colonisation de terres vers 410 Ma. Alors que les arthropodes aquatiques primitifs possèdent de nombreuses pattes, les arthropodes terrestres adoptent un mode de locomotion à six pattes qui optimise l'équilibre et la stabilité[12],[13]. La tagmatisation au niveau de la région thoracique sur laquelle est fixée trois paires de patte se traduit au niveau de la locomotion par plusieurs marches selon l'allure de l'insecte : marche longue (marche très lente avec une patte successivement au contact du sol), marche tétrapode (marche lente avec une charge importante, avec toujours quatre pattes au contact du sol), marche tripode (marche rapide avec peu de charge, avec toujours trois pattes au contact du sol)[14]. Les insectes ont le plus fréquemment une démarche tripode. Ils se tiennent sur un trépied formé par la première et la troisième patte d'un côté et la patte médiane de l'autre tandis que les trois autres pattes sont lancées vers l'avant, puis le processus s'inverse[15]. Ce mode de locomotion en « tripodes alternés » a été conservé chez les insectes alors que les vertébrés ont adopté une locomotion bipède. Une hypothèse est que que la démarche tripode permet aux insectes de se déplacer rapidement dans les terrains accidentés (en lien avec l’adhésivité des pattes qui leur permet de se déplacer à la verticale) alors que la marche bipède est plus efficace en terrain plat[16].
Les pattes présentent des morphologies variées en fonction du mode de vie des insectes qui les possèdent : pattes longues et fines pour la course, pattes courtes et trapues pour creuser, pattes en forme de rame pour nager, pattes munies de crochets pour attraper des proies, etc.
Ailes |
Les insectes modernes ont généralement deux paires d'ailes. La première paire, dite ailes antérieures (AA) se situe sur le mésothorax (T2) tandis que la deuxième, dites ailes postérieures, se trouve sur le métathorax (T3. Au sein de cette classe, on rencontre une grande variabilité de formes et de textures. D'ailleurs, la forme des ailes constitue un critère taxonomique important pour la détermination des ordres[10].
Certains insectes n'ont pas d'aile au stade adulte. Cet aptérisme est une caractéristique de plusieurs taxons d'insectes (Phthiraptera et Siphonaptera). Les insectes de l'ordre des diptères (mouche, maringouin, etc.) ont des ailes postérieures modifiées en structures appelées haltères. Les mouches sont donc considérées comme n'ayant qu'une seule paire d'ailes.
Abdomen |

L'abdomen est composé de 11 segments
L'abdomen est la partie située derrière le thorax et il comprend généralement 11 segments (A1 à A11). Ces segments possèdent une série de petits trous, appelés stigmates, qui permettent à l'insecte de respirer. Les deux principaux types sont les tergites (sur la face dorsale) et les sclérites (sur la face ventrale). Ces deux plaques sont articulées latéralement (pleure) par un repli membranaire (conjonctive) extensible, appelé repli tégumentaire pleural. Chez la plupart des insectes, les tergites sont membraneux et ils sont cachés sous les ailes au repos. Les sclérites sont généralement plus larges et ils sont bien visibles sous l'abdomen. Leur niveau de sclérification peut être très variable.
À l'intérieur, on y retrouve la majorité des organes internes (tube digestif, appareil reproducteur, etc.). L'abdomen ne porte généralement pas d'appendices pairs (ni patte, ni aile) sauf sur les segments postérieurs en position terminale : les cerques, l'ovipositeur ou encore l'aiguillon sont des appendices terminaux retrouvés chez certains insectes.
Au dernier segment (A11), l'abdomen est constitué de trois sclérites triangulaires convergeant autour des orifices anaux et génitaux. Le sclérite supérieur est appelé épiprocte tandis que les deux sclérites inférieurs sont nommés paraproctes.
Exosquelette |

Exosquelette chez les arthropodes
A : Cuticule ; B : Détails de l'épicuticule. 1 : Épicuticule ; 1a : Cément ; 1b : Cire ; 1c : Épicuticule externe ; 1d : Épicuticule interne. 2 : Exocuticule ; 3 : Endocuticule ; 2+3 : Procuticule ; 4 : Épiderme ; 5 : Membrane basale ; 6 : Cellule de l'épiderme ; 6a : Canal sécréteur de cire ; 7 : Cellule glandulaire ; 8 : Cellule trichogène ; 9 : Cellule tormogène ; 10 : Nerf ; 11 : Sensille ; 12 : Poil ; 13 : Pore glandulaire.
L'exosquelette des insectes, animaux terrestres, leur permet de résister à la pesanteur. Ce squelette externe a également pour fonction d'assurer une protection mécanique (contre les chocs, les prédateurs) et physiologique (contre la dessiccation et les infections extérieures)[17].
Le squelette externe de l'insecte est composé de deux couches : l'épicuticule et la procuticule. La première est une couche cireus qui ne contient pas de chitine. Imperméable, elle limite les pertes d'eau par transpiration. La seconde est beaucoup plus épaisse et est composée de deux couches : une externe dite exocuticule et une couche interne appelée endocuticule. L'endocuticule est résistante et flexible. Elle est faite de nombreuses couches de chitine et de protéines fibreuses entrecroisées. L'exocuticule est plus rigide et sa composition peut être très variable aux différents stades de croissance[2].
L'exosquelette peut être renforcé par un endosquelette rigidifiant l'ensemble, constitué de crêtes (épaississements locaux du tégument qui assure une protection mécanique à la tête ou renforce le thorax sollicité par les muscles puissants des ailes et des pattes), d'apophyses et d'apodèmes (invaginations tégumentaires correspondant à des prolongements internes de l'exosquelette, l'apohyse sous forme de point, l'apodème sous forme de lamelle) servant d'insertion des muscles dont l'efficacité est optimisée[18]. L'apodème est considéré comme l'équivalent du tendon des vertébrés, et est 50 fois plus rigide qu'un tendon[19]. Les insectes sont les seuls invertébrés à avoir développé la capacité de voler, cette adaptation ayant joué un rôle important dans leur succès évolutif[2]. Des muscles striés sont directement insérés sur sa face interne ou indirectement sur ces excroissances, et la disposition de ces muscles alaires permet de distinguer paléoptères et néoptères[20].
La présence d'un exosquelette, avec notamment les cires hydrophobes de l'épicuticule, protège efficacement de la dessiccation, ce qui est l'un des facteurs qui ont permis la réussite de la colonisation du milieu terrestre[21]. Mais l'exosquelette a aussi des inconvénients. Avec l'augmentation de taille, il doit devenir de plus en plus épais, de manière disproportionnée pour supporter la traction des muscles. Le cadre chitineux n'est pas aussi solide qu'un squelette osseux interne. L'internalisation du système respiratoire (constitué d'un réseau ouvert de stigmates, de trachées et de trachéoles remplies de liquide trachéolaire, dans lesquelles les gaz respiratoires se dissolvent, ce qui est nécessaire pour leur diffusion transmembranaire) protège de la dessiccation mais avec l'augmentation de taille, le rapport entre la surface intérieure des tubes et le volume du corps rend ce système respiratoire inefficace. Enfin, le confinement des muscles dans un exosquelette, les empêche de s'accroître avec leur utilisation[22]. L'exosquelette a ainsi limité la taille des insectes, ce qui favorise un rapport surface/volume élevé[23]. Ce facteur, associé à la dureté de la cuticule et au renforcement de l'endosquelette, explique que des fourmis ont la capacité de soulever et de transporter des charges de masse des centaines de fois supérieures à la leur[24],[25]. Cette petite taille a aussi rendu possible l'utilisation d'une grande variété de niches écologiques dans un environnement donné, ce qui explique qu'elle est considérée, avec le potentiel de reproduction et la diversité des insectes, comme l'élément déterminant de leur succès sur terre[26].
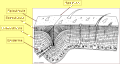
Schéma de la coupe transversale de la cuticule chez un arthropode

Exuviation d'une cigale, vue accélérée

Anatomie interne |
Système respiratoire |

Chez cette larve de Scarabée rhinocéros, la cuticule très fine à l'extrémité de l'abdomen laisse deviner un fin réseau blanc constitué des trachées et trachéoles.
Le système respiratoire des insectes est un réseau ouvert de stigmates, de trachées et de trachéoles qui assure le transfert des gaz métaboliques vers les tissus. Les stigmates des insectes, à ouverture variable, sur les côtés des segments (pleurites thoraciques et abdominaux), sont appelés spiracles[27].
Certaines larves de chironome — diptère vivant dans des milieux très faiblement oxygénés, possèdent de l'hémoglobine.
Système circulatoire |
Le milieu intérieur est constitué d'hémolymphe qui est mis en mouvement par des vaisseaux contractiles dorsaux et les mouvements musculaires généraux de l'insecte. L'appareil circulatoire est ouvert, à faible pression.
Système digestif |
L'insecte utilise son système digestif pour extraire des nutriments et d'autres substances à partir de la nourriture qu'il consomme[28]. Ces aliments sont généralement ingérés sous forme de macromolécules complexes composées de protéines, polysaccharides, lipides et d'acides nucléiques. Ces macromolécules doivent être ventilées par des réactions cataboliques pour devenir des molécules plus petites comme des acides aminés et des molécules de sucre simple. De cette manière, les cellules peuvent les assimiler.
L'appareil digestif est constitué d'un long tube clos appelé le canal alimentaire et celui-ci s'étend longitudinalement à travers le corps. Ce tube digestif régionalisé dirige unidirectionnellement la nourriture de la bouche à l'anus. Il est divisé en trois parties : stomodeum (en) (intestin antérieur) dont la fonction est le stockage et la digestion mécanique (et parfois l'aspiration), mésentéron (en) (intestin moyen) qui assure la digestion enzymatique et l'absorption des nutriments, et proctodeum (en) (intestin postérieur) où se forment les fèces. Le stomodeum et le proctodeum sont recouverts de cuticule puisqu'ils sont issus d'invaginations du tégument. En plus du tube digestif, les insectes ont également des glandes salivaires et des réservoirs salivaires. Ces structures se retrouvent dans le thorax, à côté de l'intestin antérieur[2].
Système nerveux central |
Le système nerveux central est constitué d'une double chaîne ganglionnaire ventrale, dont les ganglions les plus massifs sont antérieurs et forment le cerveau situé dans la cavité de l'exosquelette de la tête. Les trois premières paires de ganglions sont fusionnés dans le cerveau, tandis que les trois paires suivantes fusionnent pour former un ganglion sous-œsophagien qui innerve les pièces buccales[2].
Les segments thoraciques ont un ganglion placé de chaque côté du corps, donc une paire par segment. Cette disposition est également présente dans les huit premiers segments abdominaux[29]. Cette constitution peut varier, certaines blattes (blattaria) ont seulement six ganglions abdominaux. La mouche domestique (Musca domestica) a tous les ganglions fusionnés en un seul et celui-ci se retrouve dans le thorax.
Quelques insectes ont des nocicepteurs, des cellules qui détectent et transmettent des sensations de douleur[30]. Bien que la nociception ait été démontré chez les insectes, il n'y a pas de consensus sur leurs degrés de conscience à la douleur[31].
Système reproducteur |
Femelle |

Appareil reproducteur chez un papillon femelle
Chez la femelle, le système reproductif est composé d'une paire d'ovaires, des glandes accessoires, d'une ou de plusieurs spermathèques et des oviductes latéraux qui relient ces parties ensemble. Les ovaires sont constitués d'un certain nombre de tubes à œufs, appelés ovarioles. Selon les espèces, les ovaires produisent un nombre variable d'œufs.
Les glandes accessoires produisent des substances qui jouent un rôle dans la lubrification et dans l'enrobage final des œufs (ex: chorion). Ces glandes produisent également de la colle et des substances protectrices pour le revêtement des œufs (oothèque). Elles jouent aussi un rôle dans l'entretien des spermatozoïdes.
La spermathèque est un tube ou un sac qui entrepose et conserve les spermatozoïdes pendant une période (de l'accouplement à l'ovulation) très variable selon les espèces. Chez les insectes, la femelle peut gérer l'utilisation des spermatozoïdes en synchronisant leur libération de la spermathèque au moment de la ponte. Ceux-ci auront alors un accès à l'œuf passant dans l'oviducte médian[32].
Les œufs sont ensuite pondus isolés ou en masse selon l'espèce. La femelle fixera ceux-ci au substrat de ponte approprié.
Mâle |

Appareil reproducteur chez un papillon mâle
Les insectes mâles ont généralement deux testicules qui sont en suspension dans la cavité abdominale grâce aux trachées et aux corps gras. Chez les insectes plus primitifs aptérygotes, il n'y a qu'un seul testicule. Cette particularité se retrouve également chez certains lépidoptères cependant, dans leur cas, l'organe provient d'une fusion de deux testicules pendant les derniers stades de développement larvaire. Les testicules sont formés de tubes séminifères (follicules) et à l'intérieur on retrouve un sac membraneux qui contient les spermatozoïdes. Les tubes sont connectés aux canaux déférents qui servent à l'évacuation du sperme par l'intermédiaire d'un canal éjaculateur. La partie terminale de ce canal peut être sclérifiée pour former l'édéage (pénis), un organe d'intromission.
Les spermatozoïdes des insectes sont relativement longs comparativement à leur petite taille. Chez certains groupes plus ancestraux, ils sont transférés via un spermatophore. Généralement, le transfert se réalise sous forme de semence liquide par insémination directe[2].
Schéma anatomie interne |

| Anatomie de l'insecte | |
|---|---|
| A. Tête | |
| 1. antennes 2. ocelles inférieures 3. ocelle supérieure (centrale) 4. œil composé | 5. cerveau (ganglion cérébral) 31. ganglion sous-œsophagien 32. pièces buccales |
| B. Thorax | |
| 6. prothorax 7. vaisseau dorsal 8. tubes trachéaux (trompe en spirale) 9. mésothorax 10. métathorax | 27. boyaux avant (jabot, gésier) 28. ganglion thoracique 29. coxa 30. glande salivaire |
Ailes | |
| 11. première paire d'ailes | 12. seconde paire d'ailes |
Pattes | |
| 21. coussinet 22. griffes 23. tarse | 24. tibia 25. fémur 26. trochanter |
| C. Abdomen | |
| 13. boyaux médians (estomac) 14. cœur 15. ovaire (ou testicule chez le mâle) 16. boyaux arrières (intestin, rectum et anus) | 17. anus 18. vagin (ou pénis chez le mâle) 19. chaîne ganglionnaire ventrale 20. tubes de Malpighi |
Références |
Jacqueline M. C. Thomas, Les "insectes" dans la tradition orale, Peeters Publishers, 2003(lire en ligne), p. 105.
(en)Gullan, P.J.; Cranston, P.S. (2005). The Insects: An Outline of Entomology (3 ed.). Oxford: Blackwell Publishing. (ISBN 1-4051-1113-5).
(en) Richards, O. W.; Davies, R.G. (1977). Imms' General Textbook of Entomology: Volume 1: Structure, Physiology and Development Volume 2: Classification and Biology. Berlin: Springer. (ISBN 0-412-61390-5).
Nicolas Sauvion, Paul-André Calatayud, Denis Thiéry, Frédéric Marion-Poll, Interactions insectes-plantes, Editions Quae, 2013(lire en ligne), p. 107-108.
(en) Charles P. Taylor (1981). "Contribution of compound eyes and ocelli to steering of locusts in flight: I. Behavioural analysis". Journal of Experimental Biology 93 (1): 1–18.
(en) Gert Stange & Jonathon Howard (1979). "An ocellar dorsal light response in a dragonfly". Journal of Experimental Biology 83 (1): 351–355.
(en) Völkel, R; Eisner, M and Weible, K. J (June 2003). "Miniaturized imaging systems" (PDF). Microelectronic Engineering. 67–68 (1): 461–472. doi:10.1016/S0167-9317(03)00102-3.
(en) Jochen Zeil & Maha M. Al-Mutairi (1996). "Variations in the optical properties of the compound eyes of Uca lactea annulipes" (PDF). The Journal of Experimental Biology 199 (7): 1569–1577. .
(en) « Insect antennae », sur www.amentsoc.org (consulté le 25 février 2015)
(en) Johnson N. F., and C. A. Triplehorn, Borror and Delong's Introduction to the study of insects - 7th edition, Brooks Cole, 2004, 888 p. p. (ISBN 978-0030968358)
(en) « Hymenoptera: ants, bees and wasps », sur www.ento.csiro.au (consulté le 23 février 2015)
(en) Scott Richard Shaw, Planet of the Bugs. Evolution and the Rise of Insects, University of Chicago Press, 2014(lire en ligne), p. 60-63.
(en) Urless N Lanham, « Why do insects have six legs ? », Science, vol. 113, no 2945, 1951, p. 663 (DOI 10.1126/science.113.2945.663).
(en) Donald M. Wilson, « Insect walking », Annual Review of Entomology, vol. 11, 1966, p. 103-122 (DOI 10.1146/annurev.en.11.010166.000535).
Claude Nuridsany, Marie Pérennou, Microcosmos: le peuple de l'herbe, Seuil, 1998, p. 81.
(en) Pavan Ramdya, Robin Thandiackal, Raphael Cherney, Thibault Asselborn, Richard Benton, Auke Jan Ijspeert & Dario Floreano, « Climbing favours the tripod gait over alternative faster insect gaits », Nature Communications, vol. 8, no 14494, 2017.
(en) Whitney Cranshaw, Richard Redak, Bugs Rule ! An introduction to the world of insects, Princeton University Press, 2013(lire en ligne), p. 18.
(en) Whitney Cranshaw, Richard Redak, Bugs Rule ! An introduction to the world of insects, Princeton University Press, 2013, p. 15.
Lauralee Sherwood, Hillar Klandorf, Paul Yancey, Physiologie animale, De Boeck Superieur, 2016(lire en ligne), p. 350.
Michel Lamy, Les insectes et les hommes, Albin Michel, 1997, p. 135.
Jeanne Raccaud-Schoeller, Les insectes: physiologie, développement, Masson, 1980, p. 3.
Peter H Raven, Susan R Singer, Georges B Johnson, Kenneth A Mason, Jonathan B Losos, Biologie, De Boeck Superieur, 2017(lire en ligne), p. 965.
La masse d'un insecte est proportionnelle à son volume, qui est fonction du cube de ses dimensions. La force de ses muscles dépend de l'aire de leur section. Le doublement de la taille quadruple la force musculaire mais multiplie par huit la masse.
Jeanne Raccaud-Schoeller, Les insectes: physiologie, développement, Masson, 1980, p. 17.
(en) Vienny Nguyen, Blaine Lilly, Carlos Castro, « The exoskeletal structure and tensile loading behavior of an ant neck joint », Journal of Biomechanics, vol. 47, no 2, 2014, p. 497-504 (DOI 10.1016/j.jbiomech.2013.10.053).
Michel Lamy, Les insectes et les hommes, Albin Michel, 1997, p. 37.
(en)J. Buck, « Some physical aspects of insect respiration », Annu. Rev. Entomol., 7 (1962), p. 27-56
(en) "General Entomology – Digestive and Excritory system". NC state University. Retrieved 3 May 2009.
(en) Schneiderman, Howard A. (1960). "Discontinuous respiration in insects: role of the spiracles". Biol. Bull. 119 (119): 494–528.doi:10.2307/1539265. JSTOR 1539265.
(en)"Do insects feel pain? — A biological view". Cellular and Molecular Life Sciences 40: 1420–1423. 1984.doi:10.1007/BF01963580.
(en)Sømme, LS (14 January 2005). "Sentience and pain in invertebrates". Norwegian Scientific Committee for Food Safety. Retrieved 30 September 2009.
(en) Resh, Vincent H.; Ring T. Carde (2009). Encyclopedia of Insects (2 ed.). U. S. A.: Academic Press. (ISBN 0-12-374144-0).
Voir aussi |
.mw-parser-output .autres-projets ul{margin:0;padding:0}.mw-parser-output .autres-projets li{list-style-type:none;list-style-image:none;margin:0.2em 0;text-indent:0;padding-left:24px;min-height:20px;text-align:left}.mw-parser-output .autres-projets .titre{text-align:center;margin:0.2em 0}.mw-parser-output .autres-projets li a{font-style:italic}
- Anatomie des arthropodes
 Portail de l’anatomie
Portail de l’anatomie  Portail de l’entomologie
Portail de l’entomologie


