Évolution (biologie)

Arbre phylogénétique hypothétique de tous les organismes vivants, construit à partir des séquences de l'ARNr 16S. À l’origine proposé par Carl Woese (1990), enrichi par Guillaume Lecointre & Hervé Le Guyader (2006), enfin modifié par Purificación López-García & David Moreira (2008)[1], il montre l'histoire évolutive basée sur les caractères génétiques et l'analyse cladistique des trois domaines du vivant (bactéries, archées et eucaryotes).
En biologie, l’évolution est la transformation du monde vivant au cours du temps, qui se manifeste par des changements phénotypiques des organismes à travers les générations. Ces changements généralement graduels (mais pouvant être rapides ou lents) peuvent aboutir, à partir d’une seule espèce (dite « espèce-mère »), à la formation de nouvelles variétés périphériques devenant progressivement des « espèces-filles ». Inversement, la fusion de deux lignées par hybridation ou par symbiogenèse (en) entre deux populations d'espèces différentes peuvent produire une troisième espèce nouvelle. L’évolution explique la biodiversité sur Terre. L’histoire des espèces peut ainsi être pensée et représentée sous la forme d’un arbre phylogénétique et d’autres schémas et modèles, qui permettent de comprendre le phénomène de l’évolution[2].
Certains philosophes de l’Antiquité (Lucrèce, 98-54 avant notre ère, en particulier) ont approché le phénomène de l’évolution, mais ce n’est qu’à partir du XIXe siècle que des théories proposent des explications scientifiques, c’est-à-dire réfutables ou démontrables. Jean-Baptiste de Lamarck a le premier formulé une théorie scientifique transformiste fondée sur deux principes complémentaires : complexification de l'organisme et diversification adaptative.
Puis, à partir de 1859 avec la publication de L'Origine des espèces par Charles Darwin[3], le modèle darwinien de l’évolution s’est progressivement imposé dans la communauté scientifique comme celui expliquant un maximum de faits observables avec un minimum de postulats (principe de parcimonie). Darwin illustre, avec des observations détaillées, la thèse que les espèces vivantes ne sont pas des catégories fixes, mais se diversifient avec le temps, ou disparaissent[4]. Comme cause des changements qui se produisent peu à peu au sein d’une population, il propose l’idée de la sélection naturelle, équivalent naturel et spontané de la sélection artificielle pratiquée par les éleveurs d’animaux domestiques. Les espèces sont profondément conditionnées par leur milieu naturel, aujourd’hui appelé écosystème.
Toutefois, Darwin, contrairement à une croyance répandue, même à l'université, ne rejetait pas les mécanismes lamarckiens d'habituation et de transmission des caractères acquis ; il y a juste ajouté les variations spontanées et la sélection naturelle. Ce n'est qu'un an après la mort de Darwin, en 1883, qu'August Weismann a postulé la séparation des lignées germinale et somatique, ce qui implique l'impossibilité de la transmission des caractères acquis. Il ne restait donc, dans l’œuvre de Darwin plus que le mécanisme variations-sélection comme vraisemblable.
Avec la découverte de la génétique par Gregor Mendel, les modèles de l’évolution se sont peu à peu affinés[5]. Ainsi, depuis les années 1930, la théorie synthétique de l'évolution fait l’objet d’un large consensus scientifique[6]. Les recherches actuelles poursuivent l’étude des mécanismes qui permettent d’expliquer les phénomènes évolutifs[7]. Des processus découverts après 1950, comme ceux des gènes architectes, de la coévolution et de l’endosymbiose, permettent de mieux saisir les mécanismes génétiques en action, d’appréhender l’évolution des espèces les unes par rapport aux autres ou de décrire plus précisément les différents rythmes de l’évolution.
- « L'évolution biologique n'est du reste qu'un cas particulier de l'universelle évolution, car rien n'est stable : nébuleuses, étoiles, continents et mers, climats, sociétés, mœurs, religions, tout est en perpétuelle transformation »[8].
- « L'évolution signifie non seulement changement : elle implique aussi l'idée de continuité, donc celle de l'unité fondamentale de l'Univers. Alors qu'une connaissance superficielle du monde tend à le scinder en une mosaïque d'objets disparates et hétérogènes, l'œuvre de science a été constamment de ramener la diversité à une unité non de structure mais de filiation »[9].
Les logiques évolutives sont utilisées et étudiées dans des domaines aussi divers que l'agriculture, l'anthropologie, la biologie de la conservation, l'écologie, la médecine, la paléontologie, la philosophie, et la psychologie.
Sommaire
1 Histoire de la théorie de l'évolution
2 Principes généraux
3 Arguments en faveur de l'évolution
3.1 Stratégie de raisonnement
3.1.1 Utilisation des fossiles
3.2 Indices morphologiques
3.3 Indices moléculaires
3.4 Indices comportementaux
3.5 Exemple d'évolution à échelle de temps humaine : Podarcis siculus
4 Méthodes d'étude de l'évolution
4.1 Systématique
4.2 Paléobiologie
4.3 Analyse comparative des caractères
4.4 Génétique des populations
4.5 Modélisation
4.6 Expérimentation
5 Mécanismes de l'évolution
5.1 Évolution des populations
5.1.1 Apparition de nouveaux caractères
5.1.2 Variabilité des individus au sein des populations
5.1.3 Hérédité
5.1.4 Transmission des caractères acquis, une hypothèse non totalement rejetée
5.1.5 Dérive génétique
5.1.6 Sélection naturelle
5.2 Conséquences évolutives
5.2.1 Adaptation des espèces
5.2.2 Apparition et disparition des espèces
5.2.3 Sauvetage évolutif
5.2.3.1 Histoire
5.2.3.2 Modélisation
5.2.3.3 Expérimentation
5.2.3.4 Perspectives
5.3 Controverses sur les mécanismes de l'évolution
5.3.1 Evolution et complexité.
6 Évolution et sociétés humaines
6.1 Évolution et philosophie
6.2 Évolution et agriculture
6.3 Évolution et informatique
6.4 Eugénisme
6.5 Psychologie évolutionniste
6.6 Évolution et culture
6.7 Évolution et religions
6.7.1 Aspects politiques et judiciaires
7 Notes et références
8 Voir aussi
8.1 Bibliographie
8.2 Articles connexes
8.3 Liens externes
Histoire de la théorie de l'évolution |
Les hommes ont cherché l'origine de la diversité du vivant dès la période antique. L'idée d'évolution est déjà présente chez des philosophes grecs[9] et romains (Empédocle, Démocrite, Épicure, Lucrèce). Cependant, Aristote, comme beaucoup d'autres, avait une conception relativement fixiste du vivant (même si cela dépend des textes : la question ne se posant pas chez lui, il n'y apporte pas de réponse claire). Cette vision est restée prédominante dans la pensée occidentale jusqu'au XVIIIe siècle, confortée par la religion chrétienne dans laquelle toutes les espèces sont créées par Dieu au commencement du monde, déjà « parfaites » (début de la Genèse). Les grandes religions monothéistes ont diffusé cette représentation fixiste dans une vaste partie du monde. De plus ces religions confèrent à l'homme une place à part dans le vivant : il serait créé à part, à l'image de Dieu et moralement supérieur à toutes les autres espèces[9].
Durant le Moyen Âge, les débats philosophiques en Europe occidentale sont limités par la dominance du dogmatisme chrétien[10]. Les autorités religieuses condamnent fermement toute idée remettant en cause les écrits bibliques.
Dans le monde musulman, l'idée d'évolution resurgit par intermittence. Au IXe siècle, Al-Jahiz défend l'idée que non seulement les espèces évoluent au cours du temps, mais propose aussi une première théorie cherchant à expliquer cette évolution[11]. Au XIIIe siècle, le philosophe Nasir ad-Din at-Tusi soutient la sélection des meilleurs et l'adaptation des espèces à leur environnement[12]. Ces écrits se sont heurtés au dogme de la genèse et ont été oubliés pendant des siècles.
À la Renaissance des savants comme Jérôme Cardan[13], Giordano Bruno[14] et Giulio Cesare Vanini[15] remettent en cause le dogmatisme religieux, posent la question de l'origine de la vie, défendent des théories polygéniques, voire l'idée d'un ancêtre commun aux humains et aux singes. Face à l'Inquisition, certains le paieront de leur vie[16].
Au début du XVIIIe siècle, la paléontologie et la découverte de fossiles de squelettes ne ressemblant à aucun squelette d'animaux vivants[17] ébranlent les idées fixistes et font naître l'idée d'une Histoire de la nature et des espèces. Des savants redécouvrent l'idée d'évolution comme Pierre Louis Moreau de Maupertuis avec son intérêt pour l’hérédité et Georges Louis Leclerc, comte de Buffon, naturaliste passionné qui transforma le Jardin des plantes en un centre de collection et d'étude (qui deviendra le Muséum National d'Histoire Naturelle). Pour concilier ces découvertes avec les textes bibliques, Georges Cuvier expose sa théorie catastrophiste selon laquelle il y aurait eu une succession de créations divines entrecoupées d'extinctions brutales au cours des temps géologiques[18]. Il admet ainsi que les espèces terrestres n'ont pas toujours été celles observées aujourd'hui, sans pour autant accepter l'évolution des espèces (il y aurait eu plusieurs vagues successives de création), et que les 6 000 ans estimés jusque-là pour l'âge de la Terre sont trop courts pour y intégrer ces extinctions successives[19].

Jean-Baptiste de Lamarck.
La première théorie véritablement scientifique d'une évolution des espèces vivantes est avancée par le naturaliste Jean-Baptiste Lamarck. Après un long travail de classification des espèces et sur la base d'une théorie physique des êtres vivants, Lamarck développe la théorie transformiste. Il considère que les espèces peuvent se transformer selon deux principes :
- La diversification, ou spécialisation, des êtres vivants en de multiples espèces, sous l'effet des circonstances variées auxquelles ils sont confrontés dans des milieux variés et auxquelles ils s'adaptent en modifiant leur comportement ou leurs organes pour répondre à leurs besoins, généralement désigné par « l'usage et le non-usage » ;
- la complexification croissante de l'organisation des êtres vivants sous l'effet de la dynamique interne propre à leur métabolisme.
La publication, en 1809, dans Philosophie zoologique, de sa théorie transformiste entraîne de virulents débats au sein de l'Académie des sciences car elle entre en contradiction avec les idées en vigueur à l'époque et notamment le fixisme. Contrairement à une idée répandue, Lamarck n'avance aucune théorie de la transmission des caractères acquis (contrairement à ce que fera Darwin en 1868), il se contente de reprendre les idées admises sur ce point depuis Aristote.
Malgré les critiques de Cuvier, qui devient son principal opposant, les idées transformistes reçoivent une adhésion croissante à partir de 1825 et rendent les naturalistes plus réceptifs aux théories évolutionnistes[20].

Charles Darwin vers 1859, époque de la publication de "De l'origine des espèces".
Charles Darwin publie en 1859 son livre De l'origine des espèces[21] où il expose une suite d'observations très détaillées et présente le mécanisme de la sélection naturelle pour expliquer ces observations. Cette théorie évoque « la descendance avec modification » des différentes espèces. Elle considère qu'étant donné que tous les individus d'une espèce diffèrent au moins légèrement, et qu'il nait plus d'individus que le milieu ne peut en nourrir, seuls les individus les mieux adaptés à la « lutte pour la vie », c'est-à-dire à la compétition pour l'appropriation des ressources rares, parviendront à engendrer une descendance[réf. nécessaire]. Les individus ainsi sélectionnés transmettant leurs caractères à leur descendance, les espèces s'adaptent en permanence à leur milieu. Il baptise sélection naturelle cette sélection des individus les mieux adaptés en opposition à la sélection artificielle que pratiquent les agriculteurs, jardiniers et éleveurs ; cette dernière étant le socle expérimental empirique sur lequel Darwin s'appuie pour développer sa théorie.
Darwin propose dans son ouvrage de 1868, une « hypothèse de la pangenèse » qui explique la transmission des caractères acquis, mais elle sera par la suite infirmée par diverses études sur l'hérédité. August Weismann, à la fin du XIXe siècle, théorise la séparation stricte entre les cellules germinales (germen) et les cellules corporelles (soma), ce qui interdit la transmission des caractères acquis. La redécouverte des lois de Mendel à la fin du XIXe siècle et au début du XXe siècle bouleverse la compréhension des mécanismes de l'hérédité et donne naissance à la génétique. Elle est à l'origine de nouvelles méthodes dans l'étude de l'évolution, comme la génétique des populations.
Dans les années 1940, la Théorie synthétique de l'évolution, fondée entre autres par Theodosius Dobzhansky et Ernst Mayr, nait de l'articulation entre la théorie de la sélection naturelle darwinienne et de la génétique mendelienne. La découverte de l'ADN et la biologie moléculaire viennent parachever cet édifice scientifique. Depuis la biologie de l'évolution est intégrée à toutes les disciplines de la biologie et, en parallèle de son développement, contribue aussi bien à retracer l'histoire évolutive du vivant, qu'à trouver des remèdes aux maladies les plus complexes telles que le SIDA ou le cancer. Plus récemment, l'étude de l'évolution profite du développement de l'informatique et des progrès de la biologie moléculaire, notamment du séquençage du génome qui permet le développement de la phylogénie par un apport très important de données.
Principes généraux |
L'évolution est la théorie scientifique qui s'intéresse aux espèces et explique les mécanismes de leur apparition à partir d'espèces existantes ou passées. L'espèce, concept plus que réalité tangible, est le taxon de base de la systématique, bien qu'on puisse aussi parler de sous-espèces. La réalité biologique est qu'une espèce est constituée de populations dont les individus peuvent se reproduire et engendrer une descendance viable et fertile.
L'évolution du vivant commence avec l'origine de la vie il y a au moins 3,8 milliards d'années. Les premières étapes, qui ne sont pas connues précisément, ont conduit à l'apparition des trois grands groupes d'organismes actuels connus, les bactéries, les archées et les eucaryotes. L' histoire des espèces peut ainsi être pensée et représentée sous la forme d'un arbre phylogénétique.
L'évolution est constatée :
- Par la présence de variations parmi les traits héréditaires, tels que la couleur du plumage, d'une population d'individus lors des phases de reproductions avec l'apparition parfois de mutations.
- Par l'existence de divers mécanismes qui vont modifier la fréquence de certains traits héréditaires au sein de la population. Parmi ces mécanismes, la sélection naturelle désigne la différence de propagation entre les traits héréditaires causée par leur effet sur la survie et la reproduction des individus : si un certain trait héréditaire favorise les chances de survie ou la reproduction, il s'ensuit mécaniquement que la fréquence de ce trait augmente d'une génération à l'autre. Dans une population de taille finie, un trait peut également être propagé ou éliminé par le fait de fluctuations aléatoires (dérive génétique).
À l'échelle des temps géologiques, et sur le plan du phénotype, l'évolution conduit à des changements morphologiques, anatomiques, physiologiques et comportementaux des espèces. Charles Darwin a imaginé les bases de ce qui est devenu la Théorie de l'Évolution notamment en observant les ressemblances et les différences entre les différentes espèces de pinsons des différentes îles de l'archipel des Galapagos au cours de son voyage à bord du HMS Beagle. L'histoire évolutive des lémuriformes sur l'île de Madagascar est un exemple frappant illustrant la théorie de l'évolution sur un écosystème précis.
À une échelle de temps plus proche de celle que peut observer un humain, l'évolution ne se manifeste généralement qu'au sein des espèces : apparition de populations de bactéries résistantes aux antibiotiques, de populations d'insectes résistantes aux insecticides, etc. Dans certains cas toutefois, elle donne lieu à l'apparition rapide de nouvelles espèces, comme cela a par exemple sans doute été le cas pour la Pyrale du maïs (Ostrinia nubilalis), apparue en Europe à la suite de l'introduction post-colombienne de cette culture d'origine méso-américaine vraisemblablement par évolution à partir d'une espèce locale, Ostrinia scapularis[22].
Arguments en faveur de l'évolution |
Avant de considérer ces arguments, il faut rappeler qu'au sein de l'expression « théorie de l'évolution », le terme « théorie » signifie « modèle explicatif », et non pas « idée hypothétique ». Il en est de même, par exemple, avec « théorie de la gravitation » : malgré cette formulation, la réalité de la gravité ne fait pas débat.
Stratégie de raisonnement |
Si on arrive à établir un lien de parenté entre deux espèces différentes, alors cela veut dire qu'une espèce ancestrale s'est transformée en, au moins, une de ces deux espèces. Il y a donc bien eu évolution.
Un lien de parenté entre espèces fossiles ou actuelles peut être mis en évidence par le partage d'au moins un caractère homologue, c'est-à-dire provenant d'un ancêtre. Ces indices de parenté sont décelables au niveau de la morphologie, au niveau moléculaire et parfois même, pour des espèces très proches, au niveau du comportement.
Utilisation des fossiles |

Archaefructus liaoningensis, le plus ancien reste de fleur connu.
Il est en général impossible d'affirmer qu'une espèce fossile est l'ancêtre d'une espèce actuelle, car il ne sera jamais garanti que l'espèce actuelle ne s'est pas différenciée à partir d'une autre espèce proche, mais qui n'aurait pas été découverte. En effet, la conservation de restes d'espèces éteintes est un événement relativement improbable surtout pour les périodes les plus anciennes. On peut seulement estimer les liens de parenté, avec les autres espèces déjà connues, actuelles ou fossiles. Par exemple le fossile de fleur le plus ancien a été daté de 140 millions d'années. Cet organe est donc apparu sur Terre, il y a au moins 140 millions d'années. Mais d'autres espèces proches, avec des fleurs, existaient aussi certainement à cette époque. Personne n'est capable d'affirmer laquelle de ces espèces est l'ancêtre des plantes à fleur actuelles. On ne cherchera que les relations de parenté, les relations d'ancêtre à descendant ne pouvant jamais être reconstituées.
L'âge d'une espèce fossile, en revanche, indique l'âge minimum d'apparition des caractères qu'elle possède. Il est alors possible de reconstruire l'histoire de l'évolution, en plaçant sur une échelle des temps l'apparition des différents caractères. Les fossiles nous indiquent que l'ordre d'apparition des innovations évolutives est tout à fait en accord avec l'idée d'une évolution, qui dans un schéma général, part de structures simples vers des structures plus complexes. C'est aussi en accord avec une origine aquatique des êtres vivants, puisque les espèces fossiles les plus anciennes vivaient dans l'eau.
Indices morphologiques |
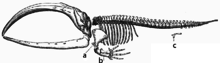
Squelette de baleine. En c figure le vestige de bassin[23].

Les « mains » des Mammifères, correspondant à l'autopode du membre chiridien ont un même plan d'organisation, ce qui traduit une homologie.
- Les baleines, animaux adaptés à la vie aquatique, gardent une trace de leurs ancêtres quadrupèdes par la présence d'os vestigiaux correspondant au bassin (ceinture pelvienne) ;
- Il y a des vestiges de pattes chez certains serpents (boas)[24] ;
- En observant l'aile d'un oiseau ou d'une chauve-souris, on retrouve aisément la structure osseuse du membre antérieur de tout tétrapode ;
- Les défenses à croissance continue des éléphants sont en fait homologues des incisives des autres mammifères, dont l'humain ;
- Les appendices masticateurs des arthropodes sont à l'origine des appendices locomoteurs réduits (il en va de même apparemment pour les Onychophores) ;
- Les membres des tétrapodes proviennent des nageoires de poissons ;
- Chez les eucaryotes, la présence d'une double membrane autour des mitochondries et la présence d'un ADN circulaire à l'intérieur de celles-ci trahissent une origine endosymbiotique procaryote. De même, dans le monde végétal, la présence d'une double membrane autour des chloroplastes et la présence d'un ADN circulaire laisse supposer l'origine cyanobactérienne de ces derniers. De surcroît, des chloroplastes à quatre membranes (plastes des chromophytes), sont les témoins d'une endosymbiose secondaire[25].
Indices moléculaires |

Animation de la double hélice d'ADN.
- Le support de l'information héréditaire est toujours l'ADN pour l'ensemble du vivant ;
- Le code génétique, code de correspondance entre l'ADN et les protéines, est quasiment le même chez tous les êtres vivants (à ne pas confondre avec le patrimoine génétique, ou génome, d'un organisme ou d'une espèce. L'expression "code génétique" étant souvent employée dans ce sens);
- Le séquençage de l'ADN. Celui-ci fait apparaître de nombreuses régions similaires, donc apparentées (gènes homologues : paralogues ou orthologues) qui codent des protéines aux fonctions ou structures différentes mais assez proches (exemple : les gènes qui codent les hémoglobines, myoglobines, etc.).
Indices comportementaux |
Chez certaines espèces de Lacertidés américains du genre Cnemidophorus, ou lézards à queue en fouet, il n'existe plus que des femelles. Ces espèces pratiquent donc une reproduction asexuée. Cependant des simulacres d'accouplements persistent : pour se reproduire une femelle monte sur une autre dans un comportement similaire à celui des espèces sexuées. Ce comportement d'origine hormonale est à mettre en relation avec une origine récente de ces espèces parthénogénétiques[26].
Exemple d'évolution à échelle de temps humaine : Podarcis siculus |

Podarcis siculus. Des lézards des ruines déposés sur l'île de Pod Mrcaru en 1971 ont évolué en 36 ans de façon à disposer d'un nouvel organe de digestion absent chez l'espèce d'origine : les valves cæcales.
Introduit en 1971 par l'équipe du professeur Eviatar Nevo sur l'île dalmate de Pod Mrcaru en mer Adriatique, le lézard des ruines (Podarcis siculus) y a été abandonné à lui-même durant plus de trois décennies, l'accès à l'île ayant été interdit par les autorités yougoslaves, puis par les conflits liés à l'éclatement de ce pays. En 2004, une équipe scientifique dirigée par Duncan Irschick et Anthony Herrel put revenir sur l'île et découvrit que Podarcis siculus avait évolué en 36 ans, soit environ trente générations, de façon très significative. Le lézard a grandi, sa mâchoire est devenue plus puissante, et surtout il a changé de régime alimentaire : d'insectivore il est devenu herbivore, et des valves cæcales sont apparues au niveau des intestins, ce qui lui permet de digérer les herbes... Cette découverte confirme, s'il en était encore besoin, que l'évolution n'est pas une théorie parmi d'autres, mais un phénomène biologique concrètement observable, et pas seulement chez les virus, les bactéries ou les espèces domestiquées[27]. Il faut cependant noter qu'il n'y a eu aucune modification de l'ADN du lézard pendant son séjour sur l'ile, ce qui revient à dire que l'information génétique était suffisante pour s'adapter à ses nouvelles conditions de vie. Il faut aussi noter qu'environ 1 % de la population des lézards des ruines possèdent des valves alors que leur régime alimentaire est resté insectivore[27]. Cet exemple ne signifie pas pour autant une évolution au sens de l'apparition d'une nouvelle espèce. Il y a une adaptation évidente de notre lézard à son nouvel environnement.
Méthodes d'étude de l'évolution |
Systématique |
Si l'on veut retranscrire les concepts en systématique, il faut considérer la théorie cladistique, selon laquelle les grades évolutifs (qui induisent une vision de l'évolution aujourd'hui obsolète[28]) ne sont plus pris en compte, en faveur des clades[29].
Paléobiologie |
La paléobiologie, étude de la vie des temps passés, permet de reconstituer l'histoire des êtres vivants. Cette histoire donne aussi des indices sur les mécanismes évolutifs en jeu dans l'évolution des espèces. La paléontologie s'occupe plus particulièrement des restes fossiles des êtres vivants. La paléogénétique, science récente, s'intéresse au matériel génétique ayant survécu jusqu'à aujourd'hui[30]. Ces deux approches sont limitées par la dégradation du matériel biologique au cours du temps. Ainsi, les informations issues des restes sont d'autant plus rares que l'être vivant concerné est ancien. De plus, certaines conditions sont plus propices que d'autres à la conservation du matériel biologique. Ainsi, les environnements anoxiques ou très froids entravent la dégradation des restes. Les restes vivants sont donc lacunaires et sont bien souvent insuffisants pour retracer l'histoire évolutive du vivant.
Analyse comparative des caractères |
Tous les êtres vivants actuels étant issus d'un même ancêtre commun, ils partagent des caractéristiques héritées de cet ancêtre. L'analyse des ressemblances entre êtres vivants donne de nombreuses informations sur leurs relations de parenté, et permet de retracer l'histoire évolutive des espèces. La phylogénie est la discipline scientifique qui cherche à retracer les relations entre êtres vivants actuels et fossiles à partir de l'analyse comparative des caractères morphologiques, physiologiques ou moléculaires. L'analyse comparative permet de retracer l'histoire évolutive des différents caractères dans les lignées du vivant. L'évolution des caractères ne suit pas nécessairement celle des espèces, certains caractères (dits convergents) peuvent être apparus plusieurs fois de manière indépendante dans différentes lignées.
L'évolution des caractères et des lignées peut être associée à des évènements géologiques ou biologiques marquant l'histoire de la Terre, ce qui permet de proposer des hypothèses sur les mécanismes à l'origine de l'évolution des espèces.
La nature des caractères pouvant être analysés est extrêmement diverse, et il peut s'agir aussi bien de caractères morphologiques (taille, forme ou volume de différentes structures), anatomiques (structure, organisation des organes), tissulaires, cellulaires ou moléculaires (séquences protéiques ou nucléiques). Ces différents caractères apportent des informations diverses et souvent complémentaires. Actuellement, les caractères moléculaires (en particulier les séquences d'ADN) sont privilégiés, du fait de leur universalité, de leur fiabilité et du faible coût des technologies associées. Ils ne peuvent cependant pas être utilisés lors de l'étude de fossiles pour lesquels seuls les caractères morphologiques sont en général informatifs.
Génétique des populations |
Modélisation |
La modélisation en biologie de l'évolution se base sur les mécanismes de l'évolution mis en évidence pour mettre en place des modèles théoriques. Ces modèles peuvent produire des résultats qui dépendent des hypothèses de départ de ce modèle, ces résultats pouvant être comparés à des données réellement observées. On peut ainsi tester la capacité du modèle à refléter la réalité, et, dans une certaine mesure, la validité de la théorie sous-jacente à ce modèle.
Les modèles dépendent souvent de paramètres, lesquels ne peuvent pas toujours être déterminés a priori. La modélisation permet de comparer les résultats du modèle et ceux de la réalité pour de nombreuses valeurs différentes de ces paramètres, et ainsi déterminer quelles sont les combinaisons de paramètres qui permettent au modèle de décrire au mieux la réalité. Ces paramètres correspondent souvent à des paramètres biologiques, et on peut ainsi estimer à partir du modèle certains paramètres biologiques difficiles à mesurer. La justesse de l'estimation de ces paramètres dépend cependant de la validité du modèle, laquelle est parfois difficile à tester.
La modélisation permet enfin de prédire certaines évolutions à venir, en utilisant les données actuelles comme données de départ du modèle.
Rappelons que le modèle est une simplification de la réalité dans un but opératoire, la prédiction ou l'explication. La théorie synthétique de l'évolution obéit à des lois et serait en partie reproductible mais elle reconnaît le rôle du hasard et de la contingence qui interviennent au niveau du gène (mutations génétiques), du génome (recombinaisons), des populations (flux et dérive génétique). Cette théorie n'est ni prévisible, ni prédictible dans la mesure où elle explique l'évolution de manière probabiliste, c'est-à-dire ni totalement déterministe, ni purement aléatoire, grâce à la sélection naturelle[31].
Expérimentation |
L'évolution expérimentale est la branche de la biologie qui étudie l'évolution par de réelles expériences, à l'inverse de l'étude comparative des caractères, qui ne fait que regarder l'état actuel des êtres vivants. Les expériences consistent généralement en l'isolement d'une ou plusieurs espèces dans un milieu biologique contrôlé. On laisse alors ces espèces évoluer pendant un certain temps, en appliquant éventuellement des changements contrôlés de conditions environnementales. On compare enfin certaines caractéristiques des espèces avant et après la période d'évolution.
L'évolution expérimentale permet non seulement d'observer l'évolution en cours, mais aussi de vérifier certaines prédictions énoncées dans le cadre de la théorie de l'évolution, et tester l'importance relative de différents mécanismes évolutifs.
L'évolution expérimentale ne peut étudier que des caractères évoluant rapidement, et se limite donc à des organismes se reproduisant rapidement, notamment des virus ou des unicellulaires, mais aussi certains organismes à génération plus longue comme la drosophile ou certains rongeurs.
Un exemple : l'expérience de Luria et Delbrück.
Mécanismes de l'évolution |
Les principaux moteurs ou facteurs de l'évolution (appelés aussi forces évolutives) sont, au niveau individuel, les mutations ponctuelles et les recombinaisons génétiques soumises au filtre de la sélection naturelle, et au niveau des populations, les flux et la dérive génétique[32].
Évolution des populations |

L'évolution des caractères dans les populations : diversité, sélection et transmission.
Parce que les individus d'une population possèdent des caractères héritables différents, et que seule une partie de ces individus accède à la reproduction, les caractères les plus adaptés à l'environnement sont préférentiellement conservés par la sélection naturelle. De plus, le hasard de la reproduction sexuée rend partiellement aléatoires les caractères qui seront transmis, par effet de dérive génétique. Ainsi, la proportion des différents caractères d'une population varie d'une génération à l'autre, conduisant à l'évolution des populations.
Apparition de nouveaux caractères |
Cela se produit par mutation et recombinaison génétique, ou remaniement chromosomique. Mais cela ne se déroule que dans un individu, pas dans l'espèce entière. Il faut, pour que ce nouveau caractère se répande, l'effet de la sélection naturelle et/ou de la dérive génétique.
De nouveaux caractères apparaissent également par symbiose lors d’endosymbiose. La fusion des partenaires initiaux peut conduire à l’émergence d’une nouvelle entité[33].
Variabilité des individus au sein des populations |
La plupart des individus d'une espèce sont uniques et différents les uns des autres[34]. Ces différences sont observables à toutes les échelles, du point de vue morphologique jusqu'à l'échelle moléculaire. Cette diversité des populations a deux origines principales : les individus sont dissemblables parce qu'ils ne possèdent pas la même information génétique et parce qu'ils ont subi des influences environnementales différentes.
La diversité génétique se manifeste par des variations locales de la séquence d'ADN, formant différents variants de la même séquence appelés allèles. Cette variabilité a plusieurs origines. Des allèles peuvent être formés spontanément par mutation de la séquence d'ADN. Par ailleurs, la reproduction sexuée contribue à la diversité génétique des populations de deux manières : d'une part, la recombinaison génétique permet de diversifier les combinaisons d'allèles réunies sur un même chromosome. D'autre part, une partie du génome de chaque parent est sélectionnée aléatoirement pour former un nouvel individu, dont le génome est par conséquent unique.
La diversité issue de l'environnement s'acquiert tout au long de l'histoire de l'individu, depuis la formation des gamètes jusqu'à sa mort. L'environnement étant unique à chaque endroit et à chaque moment, il exerce des effets uniques sur chaque individu, et ce à toutes les échelles, de la morphologie jusqu'à la biologie moléculaire. Ainsi, deux individus possédant la même information génétique (c'est par exemple le cas pour les jumeaux monozygotes ou « vrais jumeaux ») sont tout de même différents. Ils peuvent notamment avoir une organisation et une expression différente de l'information génétique.
Hérédité |
Les êtres vivants sont capables de se reproduire, transmettant ainsi une partie de leurs caractères à leurs descendants. On distingue la reproduction asexuée, ne faisant intervenir qu'un individu, de la reproduction sexuée pendant laquelle deux individus mettent en commun une partie de leur matériel génétique, formant ainsi un individu génétiquement unique.
Les caractères génétiques, c'est-à-dire l'ensemble des séquences d'acide nucléique d'un individu, ne sont pas tous transmis de la même manière lors de la reproduction asexuée, qui est une reproduction clonale, l'ensemble des séquences nucléiques est copié et l'information génétique contenue chez les deux descendants est alors identique. En revanche, lors de la reproduction sexuée, il arrive fréquemment qu'une partie seulement du matériel génétique soit transmise. Chez les Métazoaires, les chromosomes sont fréquemment associés par paire, et seul un chromosome de chaque paire et de chaque parent est transmis à l'enfant. De plus, si les parents fournissent tous les deux la moitié du contenu nucléaire, le matériel cytoplasmique est souvent fourni par un seul des deux parents (la mère chez les mammifères). Ainsi, le matériel génétique contenu dans les organites semi-autonomes, tels que les chloroplastes et les mitochondries, n'est transmis que par une partie des individus de l'espèce (les femelles chez les mammifères).
Transmission des caractères acquis, une hypothèse non totalement rejetée |
La théorie synthétique de l'évolution, paradigme dominant actuel, se fonde sur un déterminisme génétique intégral et écarte donc toute transmission héréditaire de caractères acquis au cours de la vie de l'individu. Néanmoins de plus en plus de travaux scientifiques remettent en cause ce modèle et rétablissent pour partie l'idée d'une transmission héréditaire de caractères acquis que défendait le lamarckisme[35].
Tout d'abord, certains caractères dits épigénétiques concernent la structure et l'organisation des génomes sont transmis par les parents en même temps que les molécules d'acide nucléique elles-mêmes. De plus, la mère fournit l'environnement cytoplasmique de la cellule-œuf du descendant, et transmet ainsi un certain nombre de caractéristiques cellulaires à l'enfant. Des modifications épigénétiques conservées dans la lignée germinale sont désormais décrites chez plusieurs espèces. Chez les plantes il existe une corrélation entre le niveau d'expression d'un gène et sa méthylation. Pareillement, chez les mammifères nous témoignons de la méthylation d'une séquence transposable qui est insérée à proximité d'un gène particulier. Le degré de méthylation d'un transposon pouvant enfin moduler l'expression du gène dans lequel il s'est inséré[36]. L'étude de l'épigénétique, longtemps délaissée, connait un grand essor depuis la fin du séquençage de nombreux génomes, dont celui de l'homme.
Ainsi, une étude de 2009 du MIT affirme mettre en évidence une hérédité de certains caractères acquis chez des rongeurs[37]. Par ailleurs, l’obésité serait non pas uniquement un effet direct touchant les individus atteints eux-mêmes mais également un effet transgénérationnel. Des données chez l'homme et chez l'animal semblent montrer que les effets d'une sous-alimentation subie par des individus pourraient en effet être transmis aux descendants.
Dérive génétique |

Simulation informatique de l'évolution de la fréquence d'un allèle neutre au cours du temps dans une population de 10 (en haut) ou 100 individus (en bas). Chaque courbe représente une simulation différente, les différences illustrant l'effet du hasard (dérive génétique). Les fluctuations de la fréquence de l'allèle sont plus importantes lorsque la population est de taille réduite, et la fixation (fréquence égale à 1) ou la perte (fréquence égale à 0) d'un allèle est alors plus rapide.
Lors de la reproduction sexuée, la transmission des caractères (notamment des allèles) comporte une grande part de hasard due à la recombinaison homologue, et au brassage génétique. Ainsi, on observe une variation aléatoire des fréquences alléliques d'une génération à l'autre, appelée dérive génétique. La dérive génétique génère donc une composante aléatoire dans l'évolution des populations. Ainsi, deux populations d'une même espèce n'échangeant pas de matériel génétique vont diverger jusqu'à former, si le temps d'isolement génétique est suffisant, deux espèces différentes. La dérive génétique est donc un des moteurs de la spéciation.
L'effet de la dérive génétique est particulièrement visible lorsqu'un faible nombre d'individus est à l'origine d'une population beaucoup plus nombreuse. C'est le cas lorsque se forme un goulot d'étranglement c'est-à-dire qu'une population est décimée et se reconstitue, ou lorsque quelques individus d'une population migrent pour aller coloniser un nouvel espace et former une nouvelle population (effet fondateur). Lorsqu'un tel évènement se produit, un allèle même faiblement représenté dans la population de départ peut se retrouver en forte proportion dans la population nouvellement formée sous le simple effet d'un hasard dans le tirage des individus à l'origine de la nouvelle population. Inversement, un allèle fortement représenté peut ne pas être tiré, et disparaît de la nouvelle population. Par ailleurs, la formation d'une nouvelle population à partir d'un faible nombre d'individus a pour effet d'augmenter la consanguinité dans la population et augmente le pourcentage d'homozygotie, ce qui fragilise la population.
Sélection naturelle |
Dans la très grande majorité des espèces, le nombre de cellules-œufs produit est bien plus grand que le nombre d'individus arrivant à l'âge de la maturité sexuelle et parmi ceux-ci, une partie seulement accède à la reproduction. Ainsi, seule une partie des individus formés se reproduit à la génération suivante. Il existe donc une sélection des individus perpétuant l'espèce, seuls les individus n'étant pas éliminés par les conditions environnementales pouvant se reproduire. Cette sélection a été baptisée sélection naturelle.
Comme il existe une variabilité au sein des espèces, les individus possédant des caractères différents, et qu'une partie de ces caractères sont héréditaires, les caractères permettant à l'individu de survivre et de mieux se reproduire seront préférentiellement transmis à la descendance, par rapport aux autres caractères. Ainsi la proportion des caractères au sein des espèces évolue au cours du temps.
La sélection naturelle peut prendre des formes très variées. La sélection utilitaire est une élimination des individus les moins capables de survivre et les moins féconds, alors que la sélection sexuelle conserve préférentiellement les individus les plus aptes à rencontrer un partenaire sexuel. Bien que ces sélections soient complémentaires, on observe souvent des conflits, chaque forme de sélection pouvant favoriser l'évolution d'un caractère dans un sens différent.
Il est parfois observé une sélection d'individus qui favorisent la survie ou la reproduction d'individus qui leur sont ou non apparentés, comme c'est le cas chez les insectes eusociaux ou lorsqu'un individu se sacrifie pour permettre la survie de son groupe ou de sa descendance. En sociobiologie, ces comportements altruistes s'expliquent notamment par les théories controversées de la sélection de parentèle, de la sélection de groupe et de l'altruisme réciproque. La sélection de parentèle prédit qu'il peut être plus avantageux pour un individu de favoriser beaucoup la reproduction d'un individu apparenté (donc avec lequel il partage des caractères) que de se reproduire un peu ou pas du tout, la sélection de groupe repose sur le même principe mais du point de vue du groupe et pourrait expliquer certains actes chez l'homme comme les guerres ou la xénophobie, l'altruisme réciproque se penche sur les cas d'altruisme entre individus non-apparentés et induit une contribution réciproque dont l'aide donnée en retour peut être différée dans le temps.
Enfin, la sélection artificielle n'est qu'une forme de sélection naturelle exercée par l'homme.
Conséquences évolutives |
Adaptation des espèces |
En conséquence de la sélection naturelle, les espèces conservent préférentiellement les caractères les plus adaptés à leur environnement, et y sont donc de mieux en mieux adaptées. Les pressions de sélection en jeu dans cette adaptation sont nombreuses et concernent tous les aspects de l'environnement, des contraintes physiques jusqu'aux espèces biologiques interagissantes.
L'adaptation de plusieurs espèces différentes sous l'effet des mêmes pressions environnementales peut conduire à l'apparition répétée et indépendante du même caractère adaptatif chez ces espèces, par un phénomène de convergence évolutive. Par exemple, chez les mammifères les cétacés et les siréniens ont tous deux développé des nageoires, de manière indépendante. L'évolution de ces nageoires montre une adaptation convergente à la vie aquatique.
Cependant, l'effet de la sélection naturelle est réduit par celui de la dérive génétique. Ainsi, un caractère avantageux pourra ne pas être sélectionné à cause de l'inertie donnée par la dérive. De plus, la loi de Dollo (loi sur l'irréversibilité de l'évolution) stipule qu'un caractère perdu ou abandonné au cours de l'évolution ne saurait réapparaître au sein d'une même lignée d'organismes.
L’évolution peut également se faire par mutualisme. Des relations interspécifiques s’établissent à bénéfices réciproques. Les plus abouties sont les symbioses. La théorie endosymbiotique est généralement admise aujourd’hui pour expliquer la genèse de la cellule eucaryote. L’endocytose des bactéries pourpres a rendu possible la respiration et la production d’énergie. Plus tard l’endocytose de cyanobactéries a permis la photosynthèse. Ces acquisitions ont représenté des innovations évolutives déterminantes pour l’émergence de nouvelles espèces[38]. La soudaineté de l'apparition des cellules à noyau dans les traces fossiles laisse penser que les nouvelles cellules ont été générées par un processus radicalement différent de la simple mutation ou du transfert de gènes. C'est par symbiose que des procaryotes en auraient pénétré d'autres donnant naissance aux eucaryotes. La symbiose crée sans transition de nouvelles espèces - comme les amibes bactérisées - qui n'ont pas évolué progressivement par accumulation de mutations sur une très longue durée[39].
Apparition et disparition des espèces |
L'évolution d'une population sous l'effet du hasard et des contraintes environnementales peut aboutir à la disparition de la population et éventuellement de l'espèce à laquelle elle appartient. Inversement, deux populations peuvent s'individualiser au sein d'une même espèce jusqu'à former deux espèces distinctes par un processus nommé spéciation.
Sauvetage évolutif |
| Les autres types de sauvetages | |
|
Le sauvetage évolutif est un processus théorique où une population résiste aux pressions environnementales par des changements génétiques avantageux pour échapper à l'extinction. Ces derniers peuvent avoir deux origines : une mutation qui apparaît, ou bien un allèle rare déjà présent dans la population.
On distingue le sauvetage démographique et le sauvetage génétique du sauvetage évolutif[40].
Bien que la dispersion soit un aspect clé des trois stratégies de survie de la population[41], le sauvetage évolutif est le seul qui soit guidé mécaniquement par l'évolution adaptative[42].
Histoire |
Jusqu’au XXe siècle l’évolution est comprise d’un point de vue Darwinien et Néo-Darwinien. On pense que seuls les processus écologiques expliquent les variations de populations à court terme. L’évolution n’est réservée qu’aux changements à long terme. Cependant, plusieurs observations ont montré des changements rapides dans les populations, tels que la résistance aux pesticides. Anthony David Bradshaw est le premier à soulever ce problème en 1991[43].
En 1995, Richard Gomulkiewicz et Robert Holt commencent à formaliser le concept par des modèles simples de dynamique évolutive et populationnelle dans la revue Evolution[44].
Ce concept a évolué depuis et est défini en 2012 par Andrew Gonzalez et ses collaborateurs : « le sauvetage évolutif survient lorsque l'adaptation génétique permet à une population de se rétablir des effets démographiques induits par des changements environnementaux qui, autrement, entraîneraient la disparition de l'espèce »[42]. Cette définition renforce l'idée qu’un brutal changement environnemental éloigne une population de sa niche fondamentale vers un ensemble de conditions dans lesquelles peu d'individus, voire aucun, sont capables de survivre et de se reproduire.
Modélisation |

Représentation graphique du sauvetage évolutif, la courbe démographique en U [40]Seuil de stochasticité (Stochastic threshold) : les variations génétiques en dessous de ce seuil ne sont pas déterminées par des mécanismes génétiques ou évolutifs.
Le sauvetage évolutif peut être caractérisé graphiquement par une courbe en U représentant la taille de la population au cours du temps.
Ici, on considère un changement environnemental brutal qui abaisse la fitness (valeur sélective) de la population en dessous de 1. La population ne peut donc plus se renouveler, et sa taille décroît de façon géométrique. Généralement, ce déclin mène à une extinction. Pour survivre, la population doit donc rapidement s’adapter. Chez une population en danger, l’adaptation prend une nouvelle dimension, celle de la course contre l’extinction.
- Première phase : quand l’environnement change à la génération 0, la fitness de la population descend en dessous de 1, et la taille de la population diminue.
- Seconde phase : dans le cas d’un sauvetage évolutif, la fitness rebondit au bout d’un certain moment pour dépasser 1 et la taille de la population se remet à augmenter.
- Troisième phase : la population continue ensuite de croître jusqu’à atteindre une valeur élevée et stable.
Cette courbe en U est une combinaison de deux autres courbes : le déclin géométrique des individus qui portent l’allèle sauvage et l’augmentation des individus qui portent l’allèle bénéfique[45],[40].
L’origine du sauvetage évolutif dépend du taux de mutation et de la fréquence d’allèle rare à la génération 0. Elle ne dépend pas de la fitness du nouvel allèle. En effet, les allèles rares sans avantage sélectif dans l’environnement d’origine sont susceptibles d’être perdus rapidement par stochasticité. Un sauvetage par mutation est donc plus probable. Cependant le temps requis pour l’apparition d’une nouvelle mutation bénéfique rend le sauvetage plus lent que si l’allèle existe déjà, et donc plus tardif. Ce faisant, la phase 1 de diminution de la population dure plus longtemps, et la perte de diversité est encore plus importante[45].
Expérimentation |
En plaçant une population dans un environnement stressant, on peut avoir une réaction immédiate telle que l’acclimatation (réponse physiologique pour atteindre un état d’équilibre). Sur le long terme, on observe plutôt de l’adaptation (réponse évolutive pour atteindre un état d’équilibre). Dans le cas du sauvetage évolutif, l’adaptation peut se faire sur une échelle de temps très court. L’enjeu des études expérimentales ou des observations sont donc de différencier ces deux paramètres et de les détecter[46].
| Charge génétique (genetic load) | |
Différence entre la fitness moyenne d’une population et la fitness d’une population composée entièrement d’individus avec des génotypes optimaux. Cette différence de fitness s’explique par la présence d’allèles délétères. Elle est influencée par le taux de consanguinité et la dérive génétique. |
Sous l'effet du changement environnemental, la persistance de la population dépend de la plasticité phénotypique, de la dispersion ou de l'évolution adaptative. Une expérience de Stewart & al. (2017) se concentre sur l’évolution adaptative et sa limitation par la charge génétique. Ils étudient les effets de l’évolution sur l’adaptation des petites populations lors d’un changement d’environnement grâce au modèle du coléoptère rouge de farine (Tribolium castaneum)[47]. Dans leurs expériences, les avantages de l’évolution sont contrebalancés par des processus génétiques non adaptés notamment la consanguinité et la dérive génétique. En effet, la sélection de traits avantageux réduit la diversité génétique et augmente la proportion d’homozygotes dans la population. Ils ont donc une charge génétique importante, diminuant la fitness des populations. Dans le cas d’un changement environnemental, ce phénomène associé à une petite taille de population réduit la probabilité d’un sauvetage évolutif.
Un autre exemple possible de sauvetage évolutif dans la nature est la réponse sélective des serpents noirs (Pseudechis porphyriacus) aux crapauds de canne à sucre (Bufos marinus). Ces crapauds mortellement toxiques ont été introduits en Australie en 1935, et ont rapidement provoqué la disparition de plusieurs espèces endémiques de serpents. Une étude de Ben L. Phillips et Richard Shine a révélé que les serpents noirs exposés aux crapauds avaient une préférence réduite pour ces proies, mais aussi une résistance accrue à leur toxine. Des expériences en laboratoire ont montré que ces changements n’étaient pas dû à l'apprentissage ou à l’immunité acquise après ingestion de la toxine. Ces résultats suggèrent fortement que le comportement et la physiologie du serpent noir ont évolué en réponse à la présence de crapauds, et l'ont fait rapidement : en moins de 23 générations de serpents[48].
Perspectives |
Ces études sur le sauvetage évolutif révèlent de nouvelles perspectives en biologie de la conservation. Cependant, elles permettent aussi d’expliquer certains phénomènes dans d’autre domaines d’étude, comme en médecine. En effet, la résistance aux antibiotiques de plusieurs pathogènes correspond à un sauvetage évolutif. On cherche dans ce cas à l’éviter.
Enfin, même si les populations naturelles peuvent supporter des variations environnementales intenses, elles échouent généralement à s’adapter au stress anthropique, tel que la pollution ou l’acidification. On observe alors une extinction de ces populations.
Controverses sur les mécanismes de l'évolution |
Cette section ne cite pas suffisamment ses sources (décembre 2009). Pour l'améliorer, ajoutez des références vérifiables [comment faire ?] ou le modèle {{Référence nécessaire}} sur les passages nécessitant une source. |
L'évolution et ses mécanismes sont encore largement étudiés aujourd'hui, et de nombreux points sur les mécanismes de l'évolution ne sont pas éclaircis. Certaines questions déjà soulevées par Charles Darwin n'ont d'ailleurs toujours pas de réponse certaine.
Une des grandes questions de la théorie de l'évolution est l'origine des rangs taxinomiques supérieurs à celui de l'espèce. En outre, la manière dont est apparue la majorité des 33 embranchements animaux, issus de l'explosion cambrienne, pose encore problème. Ainsi, la théorie gradualiste estime que les changements interviennent de manière progressive au cours de l'évolution, alors que la théorie des équilibres ponctués, formulée par Stephen Jay Gould et Niles Eldredge défend qu'il existe des sauts évolutifs majeurs. Selon cette théorie, le mécanisme d'évolution est tantôt accéléré tantôt ralenti, voire pratiquement nul durant de longues périodes[49]. Or au Cambrien, les paléontologues s'accordent à reconnaître des changements écologiques majeurs[50] qui pourraient selon cette théorie être à l'origine de l'apparition d'organismes appartenant aux clades actuels. De plus l'absence de fossile durant presque 100 millions d'années avant les faunes de Burgess et la rareté des sites fossilifères précambriens suggèrent l'existence de lignées fantômes précédant l'explosion cambrienne. Les formes de vie auxquelles appartiennent les animaux de Burgess n'auraient tout simplement pas été retrouvées à l'état fossile durant de longues périodes[51].
Evolution et complexité. |
Il a été longtemps admis que l'évolution s'accompagnait d'un accroissement de la complexité des êtres vivants. Cependant, cette idée, largement influencée par l'anthropocentrisme, est fortement débattue aujourd'hui[52]. La complexité n'ayant pas de définition précise à l'heure actuelle, il est difficile de vérifier une éventuelle augmentation de complexité. Par ailleurs, lorsque cette idée est admise, les origines de cette augmentation de complexité sont, elles aussi, source de controverse. En fait, tout cela a déjà été clairement expliqué par Lamarck.
Toutefois, un certain nombre d'auteurs étudient la complexité de façon formelle: Jean-Olivier Allègre, Edgar Morin et Eugene Koonin. Ce dernier, dans son livre "The Logic of Chance[53]", consacre un chapitre entier à l'origine de la complexité biologique. Il considère que « l'augmentation de la complexité est […] une tendance évolutive majeure » et que « l'émergence et l'évolution de la complexité aux niveaux du génotype et du phénotype […] (représentent) un problème central, si ce n'est le problème central en biologie » (italique dans le texte original). Plus précisément, il se demande pourquoi l'évolution n'en est pas restée au niveau des procaryotes autotrophes les plus simples, et a en fait mené à l'émergence de procaryotes complexes et, surtout, et de façon beaucoup plus frappante, des eucaryotes, avec leur génome gigantesque et régulé de façon très élaborée, de nombreux types cellulaires ; et « même capables de développer des théories mathématiques de l'évolution ».
Évolution et sociétés humaines |
Évolution et philosophie |
Le matérialisme scientifique est une forme historique et radicale de matérialisme de la seconde moitié du XIXe siècle associée au développement des sciences et à la naissance de la biologie moderne. Il défend une vision générale du monde fondée sur l'idée d'évolution et censée reposer sur les connaissances issues des sciences de la nature[54].
Aujourd'hui, on doit au physicien et philosophe Mario Bunge la réintroduction en philosophie de l'expression « matérialisme scientifique » pour désigner sa propre conception matérialiste, qu'il définit par sa proximité avec l'ensemble des sciences.[55]. Le matérialisme – qui s'affirme chez lui comme évolutionniste, organiciste, émergentiste, biologique et systémiste – s'est diversifié. Mais c'est bien la conception du matérialisme de Mario Bunge qui est aujourd'hui soutenue par Marc Silberstein, qui déclare que « le matérialisme est effectif s'il possède les attributs suivants : s'il est moniste, réaliste, scientifique, émergentiste et systémiste, réductionniste quant aux objets, aux entités et aux processus du monde mais non éliminativiste quant aux propriétés. »[56].
Marc Silberstein détaille l'unité plurielle du matérialisme « ancré dans les sciences » en reprenant les apports de Mario Bunge[57] :
- le matérialisme est à considérer comme évolutionniste, et donc, comme un bateau de Neurath.
- une doctrine ontologique.
- une méthodologie, appelée matériologie, contenant un scepticisme situé entre le doute raisonnable et le scepticisme radical.
- il existe une coévolution de la science et du matérialisme.
Un collectif autour de Muriel Gargaud et Guillaume Lecointre réactualise, de manière co-écriture multi et trans disciplinaire, le concept en soi d'évolution ainsi que l'évolution de nombreux concepts transversaux (catégorie, temps, individu, information, etc.), en lien avec l'avancée des sciences, biologiques, neurosciences, écologie, politique, de gestion, etc. et à la lumière d'une nouvelle réflexion scientifique sur le concept d'émergence[58]
Évolution et agriculture |
L'homme a su très vite utiliser la variabilité des populations à son profit : l'évolution dirigée par l'homme, ou sélection artificielle, à cause de la sélection par les éleveurs et les cultivateurs, se produit depuis des millénaires. Il avait été remarqué depuis longtemps que les animaux d'élevage héritaient, dans une certaine mesure, de caractéristiques de leurs parents et nul n'aurait songé à utiliser ses bêtes les plus malingres pour la reproduction. D'ailleurs, Darwin utilise de nombreuses observations issues de la sélection des plantes et des animaux en agriculture pour étayer ses idées. Ainsi, l'homme peut créer une sélection dite artificielle sur son environnement, volontairement pour des raisons économiques, ou involontairement via la pression de chasse, cueillette ou pêche[59].
Évolution et informatique |
L'efficacité du processus de sélection naturelle a inspiré la création d'algorithmes évolutionnistes (comme les algorithmes génétiques) en informatique. Ces algorithmes heuristiques modélisent plusieurs caractéristiques de l'évolution biologique (en particulier les mutations et les recombinaisons) pour trouver une solution satisfaisante à un problème trop complexe pour être abordé par d'autres méthodes.
Eugénisme |
La pensée évolutionniste s'est notamment propagée au sein de l'anthropologie évolutionniste au XIXe siècle. Pour les anthropologues de cette époque, l'espèce humaine ne fait qu'une, et donc, chaque société suit la même évolution, qui commence à l'état de « primitif » pour arriver jusqu'au modèle de la civilisation occidentale. Cette théorie, marquée par un très fort ethnocentrisme, du pour l'essentiel au caractère colonial et impérialiste des nations occidentales, a été très fortement remise en question par la suite. En effet, elle ne correspond pas à la réalité historique observée (les civilisations suivent des « chemins » divergents, ne poursuivent pas les mêmes « objectifs », et la civilisation occidentale, qui devrait pourtant constituer le stade ultime de l'évolution, continue pourtant à vivre de profondes mutations) et est douteuse d'un point de vue éthique (considérant la société occidentale comme l'aboutissement ultime de la civilisation). À l'inverse de ce qui était pratiqué jusqu'au milieu du XXe siècle, les approches modernes de l'anthropologie évolutionniste privilégient une méthodologie précise (confrontant des sources multiples, s'inspirant des outils d'analyse quantitative des sciences sociales, tentant de se départir de l'ethnocentrisme) et s'appuie sur des théories plus élaborées que l'évolutionnisme simpliste des débuts. Théories inspirées non seulement par la biologie de l'évolution moderne mais aussi par la modélisation mathématique et informatique et parfois enrichies par les connaissances contemporaines en psychologie.
Psychologie évolutionniste |
L'application des principes de l'évolution (notamment de concepts comme les caractères adaptatifs, la pression de sélection, etc.) en psychologie a donné naissance à un courant baptisé psychologie évolutionniste. Même si Darwin avait déjà émis l'idée que la sélection naturelle a pu façonner aussi bien des caractères anatomiques que psychologiques, cette discipline s'est véritablement formalisée au début des années 1990 dans le cadre conceptuel des sciences cognitives. Depuis, la psychologie évolutionniste est au centre d'une intense controverse scientifique qui tient à de multiples raisons : difficulté méthodologique à établir une histoire évolutive des comportements qui ne sont pas des objets matériels, résistance intellectuelle à envisager l'esprit humain comme en partie déterminé par l'évolution, utilisation simpliste et abusive des théories évolutionnistes, médiatisation et déformation auprès du grand public des problématiques scientifiques... Dans le milieu scientifique toutefois, la psychologie évolutionniste fait désormais partie des paradigmes scientifiques valides.
Évolution et culture |
Le concept d'évolution a profondément influencé la culture populaire, non seulement à travers la fascination exercée par les « mondes perdus » et les faunes préhistoriques[60] mais aussi par la généralisation de la prise de conscience que tout évolue et que l'espèce humaine n'est ni le « sommet » de l'évolution, ni une espèce « supérieure », en encore moins une espèce « prédestinée » à dominer le monde, mais une espèce-relique d'une lignée autrefois buissonnante, beaucoup plus dépendante des autres espèces qu'elles ne le sont d'elle, bref un épiphénomène de l'évolution[61].
Évolution et religions |
Les critiques de la théorie de l'évolution se répartissent en :
- Critiques idéologiques.
- Critiques religieuses (créationnisme et dessein intelligent).
Du fait, entre autres, de ses implications sur l'origine de l'humanité, l'évolution a été, et reste toujours, mal comprise et/ou, parfois, mal admise hors de la communauté scientifique. Dans les sociétés occidentales, la théorie de l'évolution se heurte à une vive opposition de la part de certains milieux religieux fondamentalistes, notamment pour son incompatibilité avec une interprétation littérale de la Bible ou du Coran. Ses détracteurs se basent sur des analyses pseudo-scientifiques ou religieuses pour contredire l'idée même d'évolution des espèces ou la théorie de la sélection naturelle.
La théorie évolutionniste est-elle compatible avec la croyance en Dieu ? En fait, Ernst Mayr dit à ce sujet : « Il me semble évident que Darwin a perdu la foi un an sinon deux, avant de formuler sa théorie de la sélection naturelle (sur laquelle il a sans doute travaillé plus de dix ans). Par conséquent, il n'est pas infondé d'avancer que la biologie et l'adhésion à la théorie de la sélection naturelle risquent de vous éloigner de Dieu »[62].
Le biologiste Richard Dawkins, dans son ouvrage Pour en finir avec Dieu (2008), pense que la sélection naturelle est « supérieure » à l'« hypothèse de Dieu » qu'il qualifie d'« improbabilité statistique », et défend l'athéisme.
Le biologiste Kenneth R. Miller (en) estime que la pensée évolutionniste n'est pas forcément incompatible avec la foi en un Dieu[63]. Pour lui les écrits de la Bible sont des métaphores.
L'évolution est encore aujourd'hui rejetée par certains milieux religieux, tenants du créationnisme, surtout protestants et musulmans.
La position de l'Église Catholique sur ce sujet est plus nuancée, tout en maintenant l'innerance de la Bible[64], « aujourd’hui, près d’un demi-siècle après la parution de l’Encyclique (Humani generis-1950), de nouvelles connaissances (la) conduisent à reconnaître dans la théorie de l’évolution plus qu’une hypothèse ». Elle déclare que Dieu est le seul créateur, qu'Il a créé le monde par amour, mais que l'esprit ne peut pas être le fruit d'une évolution de la matière[65].
Aspects politiques et judiciaires |
Les polémiques ont débordé, depuis les années 1990, le simple cadre du débat public, notamment aux États-Unis.
Dans certains États, les tenants du créationnisme ont essayé de rendre obligatoire son enseignement dans les écoles publiques, en tant que « théorie scientifique concurrente » de celle de l'évolution. Cependant ces mesures ont été déclarés anticonstitutionnelles vis-à-vis du premier amendement sur la liberté d'expression, du fait du caractère religieux de cette théorie. Devant ces tentatives, des scientifiques ont ironiquement demandé que soit aussi enseigné le pastafarisme (qui a été inventé à cette occasion).
Un nouveau concept est apparu dans la mouvance créationniste, baptisé dessein intelligent (« Intelligent Design »), qui affirme que « certaines caractéristiques de l'Univers et du monde vivant sont mieux expliquées par une cause intelligente, plutôt que par des processus aléatoires tels que la sélection naturelle »[66]. Cette thèse est présentée comme une théorie appuyée par des travaux scientifiques, et ne nie pas l'existence de tout phénomène évolutif. La justice américaine, s'appuyant sur les travaux scientifiques, a cependant jugé (voir Kitzmiller v. Dover Area School) que cette thèse était de nature religieuse et non scientifique, et que les promoteurs de l’Intelligent Design n'explicitaient pas cette « cause intelligente » afin de contourner le problème juridique et d'échapper au qualificatif religieux. D'autres groupes utilisent les arguments de l’Intelligent Design, avec diverses attributions pour la « cause intelligente », par exemple des extraterrestres.
Notes et références |
(en) Purificación López-García & David Moreira, "Tracking microbial biodiversity through molecular and genomic ecology", Research in Microbiology, Vol.159, No.1, January-February 2008, p. 67–73. DOI:10.1016/j.resmic.2007.11.019.
Corinne Fortin, Guillaume Lecointre (dir.), Alain Bénéteau (dessins), Thomas Haessig (dessins) et Dominique Visset (dessins), Guide critique de l'évolution, Paris, Belin, 2009, 571 p. (ISBN 978-2-701-14797-0, OCLC 965978019).
(en) Darwin, Charles, « Effects of the increased Use and Disuse of Parts, as controlled by Natural Selection », The Origin of Species. 6th edition, p. 108, John Murray, 1872(consulté le 28 décembre 2007).
Darwin n'utilise pas le mot évolution dans son œuvre, puisque ce terme n'est introduit que dans les années 1870. Cf. Gould (1997) : 33-37, Laurent (2001) : 17.
(en) Peter J. Bowler, The Mendelian Revolution: The Emergence of Hereditarian Concepts in Modern Science and Society, Baltimore, Johns Hopkins University Press, 1989, 1re éd. (ISBN 978-0-8018-3888-0, LCCN 89030914).
(en) Kutschera U, Niklas K, « The modern theory of biological evolution: an expanded synthesis », Naturwissenschaften, vol. 91, no 6, 2004, p. 255–76 (PMID 15241603, DOI 10.1007/s00114-004-0515-y).
Comme souvent dans le monde scientifiques, les débats y sont nombreux, comme la controverse ayant opposé Stephen Jay Gould et Richard Dawkins sur l'intérêt d'introduire la notion d'équilibres ponctués pour décrire le rythme de l'évolution.
L Cuénot et A Tétry, L'évolution biologique, Masson et Cie, 1951.
Barbara Cassin, Jean-Louis Labarrière et Gilbert Romeyer Dherbey, L'animal dans l'Antiquite, Paris, J. Vrin, 1997, 618 p. (ISBN 978-2-711-61323-6).
Jean Chaline, Quoi de neuf depuis Darwin?: la théorie de l'évolution des espèces dans tous ses états, Paris, Ellipses, 2006, 479 p. (ISBN 9782729831004 et 2729831002, lire en ligne).
Mehmet Bayrakdar (The Islamic Quarterly Third Quarter, 1983). "Al-Jahiz And the Rise of Biological Evolutionism", Londres.
Farid Alakbarli, « A 13th-Century Darwin? », Azerbaijan International, vol. 9.2, 2001, p. 48-49.
Jérôme Cardan, 1551, De Subtilitate Rerum.
En 1591 Giodano Bruno est mort brûlé pour avoir affirmé que l'humain est parent des singes, faisait valoir qu'il n'était pas crédible que les Juifs et les Éthiopiens puissent avoir le même ancêtre il y a 6000 ans, et que par conséquent, soit Dieu a créé plusieurs lignées différentes, soit les Africains sont descendants d'humains préadamiques.
Vanini et la préparation de l'esprit scientifique à l'aube du XVIIe siècle, Emile Namer, Revue d'histoire des sciences, 1972 [1].
Hans Christian Andersen (trad. Naomi Lewis, préf. Naomi Lewis, ill. Angela Barrett.), The emperor's new clothes, Cambridge, Mass, Candlewick Press, 1997(ISBN 978-0-763-60119-5), p. 25.
Laurent Dubois, [PDF] Histoire de la paléontologie, Darwin et Théorie de l'Évolution, Géopolis.fr, (page consultée le 4 juillet 2008).
Lapierre, S., « Éléments de théorie de l'évolution », sur http://www.colvir.net/prof/serge.lapierre/index.html (consulté le 19 octobre 2008).
Serge Lapierre, Éléments de théorie de l'évolution, Collège de Bois de Boulogne - Département de philosophie, (page consultée le 25 octobre 2008).
Hélène Blais, « Lamarck, genèse et enjeux du transformisme, 1770-1830 », La Revue pour l’histoire du CNRS [lire en ligne], no 7 - Novembre 2002, mis en ligne le 6 mars 2006. Consulté le 7 juillet 2008.
Darwin, C., De l'Origine des espèces, Flammarion, 1859, 1997.
(en) Vincent Calcagno, Vincent Bonhomme, Yan Thomas et Michael C. Singer, « Divergence in behaviour between the European corn borer, Ostrinia nubilalis, and its sibling species Ostrinia scapulalis: adaptation to human harvesting? », Proceedings of the Royal Society of London B: Biological Sciences, vol. 277, 7 septembre 2010, p. 2703-2709 (ISSN 0962-8452 et 1471-2954, PMID 20410041, PMCID 2982046, DOI 10.1098/rspb.2010.0433, lire en ligne)
L. Bejder, B.K. Hall, « Limbs in whales and limblessness in other vertebrates: mechanisms of evolutionary and developmental transformation and loss », Evolution & Development (en), vol. 6, no 4, nov.-dev. 2002, p. 445-58.
(en) Vestigials Organs, A Snake—With Legs ! American Museum of Natural History Site du muséum d'histoire naturelle américain.
« http://www.snv.jussieu.fr/bmedia/Chloroplaste/endosymbiose.htm »(Archive • Wikiwix • Archive.is • Google • Que faire ?) (consulté le 25 mars 2013).
Serotonergic modulation of male-like pseudocopulatory behavior in the parthenogenetic whiptail lizard, Cnemidophorus uniparens Brian George Dias et David Crews Hormones and Behavior Volume 50, Issue 3, septembre 2006, 401-409.
Voir site : [PDF](en) PNAS, vol. 105, no 12, pages 4792-4795 (25 mars 2008).
Frédéric Thomas, Michel Raymond et Thierry Lefèvre, « Encadré 1 : La diversité et la classification du vivant », dans Biologie évolutive, Louvain-la-Neuve, De Boeck Supérieur, 2016, 2e éd. (1re éd. 2010), xxiv + 965 p. [détail de l’édition] (ISBN 978-2-8073-0296-9, présentation en ligne), p. 3.
W. Hennig, Phylogenetic Systematics, Illinois University Press, 1966, traduit par D. Dwight Davis & R. Zangerl.
Eva-Maria Geigt, « L'émergence de la paléogénétique », Biofutur, no 164, février 1997, p. 28-34 (ISSN 0294-3506)
Cédric Grimoult, Le développement de la paléontologie contemporaine, Librairie Droz, 2000, p. 7.
Michel Delsol, L'évolution biologique, J. Vrin, 2002, p. 171.
Lydie Suty, Des symbioses pour mieux vivre, éditions Quæ, 2015, p. 6 à 8
(en) Maël Montévil, Matteo Mossio, Arnaud Pocheville et Giuseppe Longo, « Theoretical principles for biology: Variation », Progress in Biophysics and Molecular Biology, série From the Century of the Genome to the Century of the Organism: New Theoretical Approaches, vol. 122, no 1, 1er octobre 2016, p. 36–50 (DOI 10.1016/j.pbiomolbio.2016.08.005, lire en ligne)
« Hominidés - Théories de l'évolution en évolution », sur www.hominides.com (consulté le 25 janvier 2017)
Claudine Junien, « Obésité et diabète de type 2 : L'hypothèse de la transmission épigénétique », Cahiers de nutrition et de diététique, vol. 37, no 4, 2002, p. 261-272 (ISSN 0007-9960, lire en ligne)
(en) Emily Singer, « A Comeback for Lamarckian Evolution? », MIT Technology Review, 4 février 2009(lire en ligne)
Lydie Suty, déjà cité
Lynn Margulis et Dorion Sagan, L'univers bactériel, Albin Michel, 1989, p.122, 129
(en) S.M Carlson, « Evolutionary rescue in a changing world », Trends in Ecology & Evolution, vol 29, No 9., septembre 2014
(en) Bell G., « Evolutionary rescue and the limits of adaptation », Philosophical Transactions of the Royal Society B: Biological Sciences, 10 décembre 2017
(en) Gonzalez A, Ronce O, Ferriere R, Hochberg ME, « Evolutionary rescue: an emerging focus at the intersection between ecology and evolution. », Philosophical Transactions of the Royal Society B: Biological Sciences., février 2012
(en) Bradshaw, A. D., « Genostasis and the limits to evolution », Philosophical Transactions of the Royal Society of London. Series B, 1991
(en) R. Gomulkiewicz, R. Holt, « When does natural selection save a population from extinction? », Evolution, 1995
(en) Orr HA, Unckless RL, « The Population Genetics of Evolutionary Rescue », PLOS Genetics, 2014
(en) G. Bell, S. Collins, « Adaptation, extinction and global change », Evolutionary Applications, 2008
(en) Gavin S. Stewart et al., « The power of evolutionary rescue is contrained by genetic load », Evolutionary Applications, 2017
(en) B.L Phillips, R. Shine, « An invasive species induces rapid adaptive change in a native predator: cane toads and black snakes in Australia », Proceedings: Biological Sciences, 2006
Stephen Jay Gould et Marcel Blanc, La structure de la théorie de l'évolution, Gallimard, 2006(ISBN 2070766810 et 9782070766819)
Landing, E.; MacGabhann, B. �N. A. (2009). "First evidence for Cambrian glaciation provided by sections in Avalonian New Brunswick and Ireland: Additional data for Avalon–Gondwana separation by the earliest Palaeozoic". Palaeogeography, Palaeoclimatology, Palaeoecology
Hors-série Science et vie, Mondes disparus.
Cette idée a notamment été critiquée par Stephen Jay Gould dans Stephen Gould (trad. Christian Jeanmougin), L'éventail du vivant : le mythe du progrès [« The Spread of Excellence From Plato to Darwin »], Paris, Editions du Seuil, coll. « Sciences » (no S143), 1997(ISBN 978-2-020-49093-1, OCLC 47826010)
Koonin, Eugene V., The logic of chance : the nature and origin of biological evolution, Pearson Education, 2012(ISBN 9780132542494, 0132542498 et 0133381064, OCLC 711043216, lire en ligne)
Pascal Charbonnat, Histoire des philosophies matérialistes, Paris, Kimé, 2013, 706 p. (ISBN 9782841746224, lire en ligne).
Collectif sous la direction de Marc Silberstein, Matériaux philosophiques et scientifiques pour un matérialisme contemporain, t. 1, Paris, Éditions Matériologiques, 2013, 1017 p. (ISBN 9782919694259), p. 97 à 100
Collectif, Intrusions spiritualistes et impostures intellectuelles en sciences, Paris, Éditions Syllepse, coll. « Matériologiques », 2001, 399 p. (ISBN 9782913165670, lire en ligne), p. 13 à 15
Article :L'unité plurielle du matérialisme de Marc Silberstein et Collectif, 2013, 1er tome, p. 7 à 27.
Collectif, L'évolution : de l'univers aux sociétés : Objets et concepts, Paris, Éditions Matériologiques, coll. « Science & philosophie », 2015, 502 p. (ISBN 9782373610321, lire en ligne).
Chris T. Darimonta et al. ; Human predators outpace other agents of trait change in the wild ; Ed : Gretchen C. Daily, Stanford University, Stanford, CA, PNAS, approuvé le 21 novembre 2008 (reçu pour relecture le 15 septembre 2008) (« Lire l'article »(Archive • Wikiwix • Archive.is • Google • Que faire ?) (consulté le 25 mars 2013))
L'évolution vue par Walt Disney dans son film Fantasia sur [2] : on y voit entre autres les principales espèces de dinosaures du Mésozoïque mélangées sur la musique du Sacre du printemps d'Igor Stravinsky.
Guillaume Lecointre (dir.) et Corinne Fortin (dir.), Guide critique de l'évolution, Paris, Belin, 2009, 571 p. (ISBN 2-701-14797-2, OCLC 965978019)
Neil A Campbell, Biologie, Ed. du Renouveau pédagogique ; De Boeck Université, 1995(ISBN 9782761306539 et 9782804120849), p. 417
Kenneth Miller À la recherche du Dieu de Darwin, édition Sciences et quête de sens, 2000
« Des sources du Pentateuque et de l'historicité de Genèse 1-11 - Pontificia Commissione Biblica », sur www.vatican.va (consulté le 25 janvier 2017)
Intervention du Pape Jean-Paul II devant l'Académie Pontificale des Sciences le 22 octobre 1996.
« The theory of intelligent design [...] holds that certain features of the universe and of living things are best explained by an intelligent cause rather than an undirected process such as natural selection » — Intelligent Design Network, Inc.
Voir aussi |
Bibliographie |
Lorsqu'il y a deux dates, la première est celle de la première parution, dans la langue d'origine.
- Brondex, F. (1999) Évolution : synthèse des faits et théories, Dunod.
Buican, D. (1989) La Révolution de l'évolution, Presses universitaires de France.
Buican, D. (1997) L'Évolution et les théories évolutionnistes, Éditions Masson.
Buican, D. (2008) L'odyssée de l'évolution, Ellipses
Chapouthier, G. (2001) L'homme, ce singe en mosaïque, Éditions Odile Jacob
- Combes, C. (2006) Darwin, dessine-moi les hommes
Darwin, C. (1997, éd. or. 1859) L'Origine des espèces, Flammarion.
Charles Darwin. Origines - Lettres choisies 1828-1859 (2009), introduction et édition française dirigée par Dominique Lecourt, préface S. J. Gould, éditions Bayard, (ISBN 978-2-227-47843-5).- David, P. & Samadi, S. (2000) La Théorie de l'évolution, Flammarion.
Dawkins, R. (1982) The Extended Phenotype, Oxford University Press.
Dawkins, R. (1986, 1989) L'Horloger aveugle, Éditions Robert Laffont.
Dawkins, R. (1996) Climbing Mount Improbable, Norton (anglophone).
Dawkins, R. (1976, 1996) Le Gène égoïste, Odile Jacob.- Dennett, D. (2000) Darwin est-il dangereux ?, Odile Jacob.
- Michael Denton, 1985, Evolution : A theory in crisis, (Évolution - Une théorie en crise), Flammarion.
- Devillers, C. & Tintant, H. (1996) Questions sur la théorie de l'évolution, PUF.
- Dorléans, P. (2003) Il était une fois l'évolution, Ellipses.
- Futuyma, D.J (1997) Evolutionary Biology, Sinauer Associates.
Gould, S. J. (1982) Le Pouce du panda, Éditions Grasset.
Gould, S. J. (1991) La Vie est belle, Éditions du Seuil.
Gould, S. J. (1997) L'Éventail du vivant, Le Seuil.
Gould, S. J. (1997). Darwin et les grandes énigmes de la vie. Réflexions sur l'histoire naturelle. 1, S 43, Seuil (Paris), collection Point Science : 311 p.
Gould, S. J. (2000) Et Dieu dit : Que Darwin soit ! : Science et religion, enfin la paix ?, préface de D. Lecourt, Le Seuil.
Gould, S. J. (2002) The structure of evolutionary theory, Harvard University Press (anglophone).
Grassé P. P. (1973) L'Évolution du vivant, matériaux pour une théorie transformiste, Albin Michel.- Grimoult, C. (2000) Histoire de l'évolutionnisme contemporain en France (1945-1995), Genève, Droz.
- Grimoult, C.(2001) L'évolution biologique en France. Une révolution scientifique, politique et culturelle, Genève, Droz.
- Guillo, D. (2009) Ni Dieu ni Darwin, les Français et la théorie de l'évolution, Ellipses
Jacob, F. (1981) Le Jeu des possibles, Fayard.
Kropotkine, P. (2001, éd. or. 1902) L'entraide : un facteur de l'évolution, Ecosociété.- Laurent, G. (2001). La Naissance du transformisme. Lamarck entre Linné et Darwin, Vuibert (Paris) et ADAPT (Paris) : 151 p. (ISBN 978-2-7117-5348-2)
Guillaume Lecointre, L'évolution - Question d'actualité ?, Paris, Quae, 2014, 108 p. (ISBN 9782759222513, lire en ligne).
Dominique Lecourt, (1992, 3e éd. « Quadrige » 1998), L’Amérique entre la Bible et Darwin, suivi de Intelligent design : science, morale et politique, Presses universitaires de France.
Lecourt, D., dir. (1999, 4e réed. « Quadrige » 2006), Dictionnaire d’histoire et philosophie des sciences, PUF.
Le Guyader, H., dir. (1998) L'évolution, Belin/Pour la Science.
Le Guyader, H. et Lecointre, G., Classification phylogénétique du vivant, Éditions Belin (Paris) : 560 p. (ISBN 978-2-7011-4273-9)
- Lehman, J.-P. (1973) Les preuves paléontologiques de l'évolution, PUF.
Lodé, T. (2006) La guerre des sexes chez les animaux, Odile Jacob, Paris. (ISBN 978-2-7381-1901-8)
- Marchand, D. (2002) Les merveilles de l'évolution, P.U. Dijon.
Mayr, E. (1989) The Growth of Biological Thought: Diversity, Evolution and Inheritance, Ed. Cambridge, Harvard University Press — traduit en français sous le titre de Histoire de la biologie. Diversité, évolution et hérédité, Fayard (1989) : 894 p. (ISBN 978-2-213-01894-2).
Mayr, E. (2004) What makes biology unique? Considerations on the Autonomy of a Scientific Discipline, Ed. New York, Cambridge University Press — traduit en français sous le titre de Ernst Après Darwin. La biologie, une science pas comme les autres, Dunod (2006) : 237 p. (ISBN 978-2-10-049560-3).
Pichot, A. (1993) Histoire de la notion de vie, éd. Gallimard, coll. TEL.- Ridley, M. (1998) Évolution Biologique, Éd. De Boeck (traduction française).
Tort, P. (1996) Dictionnaire du darwinisme et de l’évolution, Ed. Paris, PUF, 3 vol., 5 000 p. Ouvrage couronné par l’Académie des Sciences.- James A. Shapiro (2011). Evolution: A View from the 21st Century. FT Press. (ISBN 9780132780933)
- Wright, R. (1995) L’Animal Moral, Michalon.
- Zimmer, K. (2001) Evolution: the triumph of an idea, Harper Collins (anglophone)
- Collectifs
- Collectif, L'évolution : de l'univers aux sociétés : Objets et concepts, Paris, Éditions Matériologiques, coll. « Science & philosophie », 2015, 502 p. (ISBN 9782373610321, lire en ligne)
.mw-parser-output .autres-projets ul{margin:0;padding:0}.mw-parser-output .autres-projets li{list-style-type:none;list-style-image:none;margin:0.2em 0;text-indent:0;padding-left:24px;min-height:20px;text-align:left}.mw-parser-output .autres-projets .titre{text-align:center;margin:0.2em 0}.mw-parser-output .autres-projets li a{font-style:italic}
Articles connexes |
- Adaptation
- Arbre phylogénétique
- Biodiversité
- Biologie de l'évolution
- Blessure narcissique
- Charles Darwin
- Cladistique
- Classification phylogénétique du vivant
- Coévolution
- Conflit sexuel
- Conséquences évolutives de la compétition
- Créationnisme
- Débat entre Huxley et Wilberforce
- Débat sur l'hippocampe
- Évolution moléculaire
- Évolution dirigée
- Evolution de l'allocation à la reproduction
- Eukaryota (classification phylogénétique)
- Équilibre ponctué
- Ève mitochondriale
- Évolution convergente
- Évolution insulaire
- Exaptation
- Fixisme
- Gène égoïste
- Génération spontanée
- Géographie physique
- Histoire de la pensée évolutionniste
- Historique des critiques des théories de l'évolution
- Innovation évolutive
- Jean-Baptiste de Lamarck
- Mémétique
- Microévolution et macroévolution
- Naissance du vivant
- Nouveauté évolutive
- Panspermie
- Photo-guide taxinomique du monde animal
- Photo-guide taxinomique du monde végétal
- Prédéterminisme
- Robustesse
- Sélection naturelle
- Sélection de parentèle
- Sensibilité des plantes
- Spéciation
- Systématique évolutionniste
- Taxinomie
- Théorie du handicap
- Théorie synthétique de l'évolution
- Théorie synergique de l'évolution
- Évolution du mimétisme chez les insectes
- William Donald Hamilton
Liens externes |
(fr) Évolution. De l'origine de la vie aux origines de l'homme, dossier SagaScience du CNRS (France).- Sélection de sites web sur la systématique et l’évolution dans le répertoire encyclopédique : Les Signets de la Bibliothèque nationale de France
- Effervesciences (CINAPS Télévision) : Darwin aujourd'hui (avec Guillaume Lecointre)
André Pichot, Sur la notion d'évolution en biologie, 2007.
(en) L'évolution résumée en 60 secondes
 Portail de l’évolution
Portail de l’évolution


